 2019, Vol. 38
2019, Vol. 38扩展功能
文章信息
- 陈雪, 张塔星, 罗概, 吴永杰, 冉江洪, 陈明华, 李良, 龚亿华
- CHEN Xue, ZHANG Taxing, LUO Gai, WU Yongjie, RAN Jianghong, CHEN Minghua, LI Liang, GONG Yihua
- 四川美姑大风顶国家级自然保护区鸟类物种及垂直多样性研究
- Composition and Vertical Distribution Pattern of Birds Species in the Meigu Dafengding National Nature Reserve, Sichuan
- 四川动物, 2019, 38(4): 445-451
- Sichuan Journal of Zoology, 2019, 38(4): 445-451
- 10.11984/j.issn.1000-7083.20190024
-
文章历史
- 收稿日期: 2019-01-08
- 接受日期: 2019-06-06






2. 四川美姑大风顶国家级自然保护区管理局, 四川美姑 616450
2. Meigu Dafengding National Nature Reserve Administration Bureau, Meigu, Sichuan Province 616450, China
鸟类多样性研究一直是群落生态学研究的重要领域,对于整个群落生态学概念的形成和发展起着非常重要的作用(崔鹏,邓文洪,2007),鸟类多样性可直接反映栖息地的适宜性、生态系统健康与生物多样性状况、人类活动对生态系统的干扰程度以及区域生态环境的质量等(斯幸峰,丁平,2011)。鸟类多样性的垂直分布格局是进化过程和生态过程共同作用的结果,并且在不同地区和气候条件下具有不同模式(Price et al., 2014;Pontarp & Wiens,2017;Quintero & Jetz,2018),了解不同地区,特别是生物多样性热点地区和未充分或者缺少调查地区的鸟类多样性垂直分布模式及潜在维持机制对于生物多样性的保护至关重要(Acharya et al., 2011)。
四川美姑大风顶国家级自然保护区地处青藏高原东南部的横断山脉与四川盆地的西南边缘交汇处,位于凉山山系的核心区域,是以大熊猫等珍稀濒危野生动物及其栖息地为主要保护对象的保护区。保护区一直未开展系统的鸟类资源调查,保护区管理局对区内的鸟类多样性还缺乏足够的了解,这限制了保护区保护管理策略的制订和保护成效的提升。整个凉山山系鸟类的研究也很缺乏,除施白南和赵尔宓(1982)记录了凉山山系部分县的鸟类种类,李桂垣等(1984)总结分析了凉山彝族自治州的鸟类多样性外,近30年来,基本没有凉山山系鸟类多样性的文献报道,导致我们对凉山山系这一重要生物多样性保护地区的鸟类多样性及其变化情况缺乏了解。本文根据实地调查并综合历史文献资料,对保护区的鸟类名录进行了较全面的整理与归纳,以期为保护区的鸟类资源保护以及保护策略制订提供基础资料,进一步充实凉山山系的鸟类分布数据;同时在保护区开展鸟类垂直多样性的研究,也能够为小尺度下鸟类多样性垂直分布格局提供新的研究案例。
1 研究地概况四川美姑大风顶国家级自然保护区位于四川省凉山彝族自治州美姑县东北部,属青藏高原的东南缘,102°52′~103°20′E,28°30′~28°50′N,最高海拔3 998 m,最低海拔1 356 m,总面积50 655 hm2。保护区属于中亚热带季风湿润气候,地处低纬度高海拔山区,具有低纬度高原性气候特点,气温年差较小,日差较大。根据县内气象划分四季:3—5月为春季,6—8月为夏季,9—10月为秋季,11月至翌年2月为冬季。年平均气温10.2 ℃,年平均降水量1 089 mm,相对湿度80%,无霜期280~230 d。区内地形复杂多样,由低海拔到高海拔植被的垂直带谱分布表现明显:1 800 m以下为常绿阔叶林带,1 800~2 200 m为常绿与落叶、阔叶混交林带,2 200~2 500 m为针阔叶混交林带,2 500~3 500 m为亚高山针叶林带,3 500 m以上为高山灌丛草甸带(宋昭彬等,2004)。因人类活动和历史原因,海拔1 800~3 200 m还分布有人工种植的针叶林,主要包括日本落叶松Larix kaempferi、云南松Pinus yunnanensis、川滇冷杉Abies forrestii等。
2 研究方法 2.1 调查方法2016年12月—2018年6月,采用样带法和样点法在保护区及周边海拔1 800~3 800 m对鸟类共展开了7次野外调查:2016年12月、2017年4月和6月采用样带法对保护区鸟类多样性开展调查,共计52条样带;2017年6月、9月、12月、2018年3月和6月采用样点法对森林鸟类开展系统的重复调查,每次调查周期约1个月。7次野外调查时间总计174 d。样带和样点位置详见图 1。

|
| 图 1 美姑大风顶国家级自然保护区位置及野外样点 Fig. 1 Location and field sampling point of the Meigu Dafengding National Nature Reserve |
| |
样带法采用典型样带调查,样带涵盖保护区的各类生境和典型区域,长度与各类生境面积成正比,即各植被类型的抽样强度基本与区域的背景值相当。样带调查时以1~3 km·h-1的速度行进,使用双筒望远镜观察记录样线两侧和前方鸟类,并辅以相机拍照和鸣声辨别,记录鸟类物种、个体数量、行为、栖息生境及海拔等。考虑取样的充分性,在视线受限的森林生境采用样点法观测,共设置不同海拔的36个样点,半径50 m,样点之间距离大于300 m。每天调查8 h(07: 00—11: 00、14: 00—18: 00),使用定时点数统计法计数,记录所有出现鸟类的种类、数量、活动类型(鸣叫、鸣唱、飞行、休息、觅食和筑巢)、生境类型、海拔和是否混群等,并估测目标鸟类与调查人员之间的距离。中大雨、大风及大雾天气不调查。
2.2 分析方法鸟类分类系统和特有种参照《中国鸟类分类与分布名录(第三版)》(郑光美,2017),居留型和区系分别参照《四川鸟类鉴定手册》(张俊范,1996)和《中国动物地理》(张荣祖,2011)。由于部分海拔未设置调查样点,鸟类丰富度的垂直分布格局分析仅使用样带调查法的数据,分为5个海拔段(图 3),并采用插值物种丰富度及Mao's tau丰富度进行研究。插值物种丰富度为某一物种若分别在较低海拔和较高海拔地区均被发现,那么可认为该物种在2个海拔之间的地段也有分布(Wu et al., 2013;吴永杰等,2017);Mao's tau丰富度是基于不同海拔段样线上的观测物种存在或缺失矩阵计算出的物种丰富度(Colwell,2004)。为了保证不同海拔段的抽样强度相等,以最小抽样强度海拔段(3 400~3 800 m)的抽样强度为标准来计算其他海拔段的Mao's tau丰富度。

|
| 图 3 美姑大风顶国家级自然保护区鸟类的物种丰富度垂直分布格局 Fig. 3 Elevational pattern of birds richness in the Meigu Dafengding National Nature Reserve |
| |
不同生境间的鸟类群落结构相似性采用Jac card相似性指数(IsJ),IsJ=C(A,B)/,其中,C(A,B)是生境A和生境B均拥有的物种数,N(A)+N(B)为生境A和生境B的总物种数(McCune et al., 2002)。相关分析使用Pearson法,Mao's tau丰富度的计算与分析在Past 3中完成,其他数据分析在SPSS和R 3.5.1中完成。
3 结果 3.1 鸟类组成2016—2018年野外调查记录保护区鸟类177种,隶属13目44科;结合历史资料(施白南等,1982;内部资料),共记录鸟类17目56科268种(附录1)。其中,雀形目Passeriformes鸟类37科200种,占鸟类总种数的74.6%,非雀形目20科68种,占25.4%。
国家Ⅰ级重点保护野生鸟类2种;国家Ⅱ级重点保护野生鸟类16种;列入濒危野生动植物种国际贸易公约(CITES)附录Ⅰ1种,附录Ⅱ 12种,附录Ⅲ 1种;列入世界自然保护联盟(IUCN)濒危物种红色名录濒危(EN)1种,易危(VU)3种,近危(NT)2种;中国特有种14种。
3.2 鸟类居留型和区系特征保护区分布的268种鸟类中,留鸟146种(54.5%),夏候鸟91种(34.0%),冬候鸟18种(6.7%),旅鸟13种(4.9%),以繁殖鸟为主。在237种繁殖鸟中,东洋界160种(67.5%),古北界54种(22.8%),广布种23种(9.7%)。
3.3 生境分布将保护区鸟类的栖息微生境主要分为针叶林、针阔混交林、阔叶林、灌丛及灌草丛、高山灌丛草甸和水域(包括水体及岸区)6种,统计实际调查的177种鸟类的微生境分布,针叶林和阔叶林2种生境类型中的鸟类物种数最多,水域和高山灌丛草甸中的最少(表 1)。
| 针叶林 | 针阔混交林 | 阔叶林 | 灌丛及灌草丛 | 高山灌丛草甸 | 水域 | |
| 物种数 | 142 | 100 | 129 | 74 | 28 | 18 |
| 占比/% | 80.2 | 56.5 | 72.9 | 41.8 | 15.8 | 10.2 |
不同生境的鸟类群落结构相似性分析结果显示:相似性高的分别是针叶林与阔叶林、针叶林与针阔混交林、针阔混交林与阔叶林,IsJ分别为59.8%、55.1%、48.7%,即森林生境内的鸟类群落结构相似性最高,而水域与其他生境的最低(表 2)。
| 生境类型 | 针叶林 | 针阔混交林 | 阔叶林 | 灌丛 | 高山草甸 | 水域 |
| 针叶林 | 86 | 101 | 61 | 22 | 12 | |
| 针阔混交林 | 55.1% | 75 | 50 | 17 | 12 | |
| 阔叶林 | 59.8% | 48.7% | 57 | 19 | 9 | |
| 灌丛 | 39.4% | 40.3% | 39.0% | 17 | 8 | |
| 高山草甸 | 14.9% | 15.3% | 13.8% | 20.0% | 2 | |
| 水域 | 8.1% | 11.3% | 6.5% | 9.5% | 4.5% | |
| 注:右上表示对应2个生境中均存在的鸟种数;左下表示对应2个生境的Jaccard相似性指数 Note:the upper right data indicate the number of birds recorded in both habitats;the lower left data indicate the Jaccard similarity index for birds between 2 habitats | ||||||
在177种实际观察的鸟类中,候鸟76种,且夏季(57种)>春季(46种)>秋季(30种)>冬季(14种),夏季最多,冬季最少。夏候鸟在各季节均占最高比例,其迁入迁出可能是导致保护区不同季节鸟类群落组成变化的主要原因。冬候鸟和旅鸟在相应季节较少,但全年均有观测记录(图 2)。

|
| 图 2 美姑大风顶国家级自然保护区候鸟的季节变动 Fig. 2 Seasonal changes of migratory birds in the Meigu Dafengding National Nature Reserve |
| |
插值物种丰富度、观测值和Mao's tau丰富度均表明保护区鸟类在海拔段2 600~3000 m的物种丰富度最高,在海拔段3 400~3 800 m最低,鸟类垂直分布整体呈中峰模式。插值丰富度和Mao's tau丰富度有着相同的垂直分布模式(r=0.928,P=0.023),中低海拔段(1 800~2200 m和2 200~ 2 600 m)的物种丰富度比中高海拔段(3 000~3 400 m和3 400~3 800 m的)更高(图 3)。
4 讨论本次调查野外实际调查到177种,占保护区鸟类总物种数的66.0%,另有91种未记录到,还需要保护区进一步调查和确认。在保护区分布的16种国家重点保护鸟类中,仅调查确认到8种。未发现的国家Ⅰ级重点保护鸟类四川山鹧鸪Arborophila rufipectus,仅李操等(2003)在1998年通过访问调查有分布,未见实体;虽然戴波等(2014)通过卫片解译,发现区内存在大片潜在适宜生境,与保护区相邻同属于凉山山系的马边彝族自治县也有实体分布,但我们2年的调查没有记录到该物种,是否存在实体尚待进一步研究。国家Ⅱ级重点保护野生鸟类大紫胸鹦鹉Psittacula derbiana由郑作新等(1962)于1960年在美姑县采得标本,但在2004年保护区的第一次科学考察中没有发现其痕迹,保护区的长期监测也未记录到该物种,通过社区访问也无人在野外发现过该物种,估计在保护区已经绝迹。
由于居留型的划分主要是根据已有调查记录进行划分,但鸟类的居留型并非一成不变,其受调查资料的积累和气候的影响较大。如普通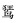
保护区针叶林中的鸟类物种数最多,这可能主要由面积效应造成,即针叶林面积最大,较大的面积能够容纳更多的物种(Rahbek,2005)。同时,生境中的植被特征也被认为是决定鸟类群落多样性最重要的因素之一(MacGregor-Fors & Schondube,2011)。保护区鸟类物种数在针叶林、针阔混交林以及阔叶林3种森林生境中最高,而在其他生境中较低。由于森林生境的空间异质性高,内部结构复杂,拥有更多的小生境及小气候条件,进而满足更多具有不同生态位的鸟类需要(Mentil et al., 2018),因此,森林比生境结构相对同质化的草甸及灌丛和灌草丛能支持更高的鸟类多样性。在野外调查中,金色林鸲Tarsiger chrysaeus、三趾啄木鸟Picoides tridactylus等森林依赖种仅在森林生境被发现,小云雀Alauda gulgula、灰眉岩鹀Emberiza godlewskii等开阔生境种仅在草甸和灌丛生境被发现,研究结果也显示森林与开阔生境(草甸、灌丛和灌草)间的相似度比3种开阔生境之间的相似度低,这可能是由于不同类群鸟类在其适应的生境内进化,导致其生物学特征和生境类型密切相关,越相似的生境可支持物种组成更相似的鸟类群落(Zhang et al., 2011),不同生境类型对于维持不同类群的鸟类以及整个鸟类群落的多样性十分重要(Ricklefs,1991)。
本研究还发现保护区的鸟类丰富度具有显著的垂直格局。物种丰富度垂直格局通常分为4类:峰值位于中海拔的中峰模式、单调递减模式、低海拔平台然后随海拔升高递减模式和峰值偏向低海拔的单峰分布模式(McCain,2009)。本研究结果属于中峰模式,鸟类丰富度的峰值出现在2 600~3 000 m的中海拔段,这一类型的分布格局与多数研究结果一致(Rahbek,2005;吴永杰,雷富民,2013;梁丹等,2015)。梁丹等(2015)调查了高黎贡山中段海拔的鸟类,发现鸟类丰富度整体呈中峰模式,但与韩联宪等(1996)报道的丰富度峰值出现在更低的海拔不同,这可能与两者研究所涉及的海拔跨度不同有关。美姑大风顶国家级自然保护区由于整体处于中高海拔的山脊,保护区外的低海拔均分布有农田和民居,这限制了本研究对整个海拔梯度的采样。虽然保护区鸟类垂直分布格局总体呈中峰模式,但在更大海拔梯度下凉山山系的鸟类垂直分布格局及影响机制还需进一步加强保护区邻近地区的研究。
附录 Supplementary material附录1 四川美姑大风顶国家级自然保护区鸟类名录
Appendix 1 The checklist of the birds in the Meigu Dafengding National Nature Reserve,Sichuan(http://www.scdwzz.com/Articles/fujian/20190024-1.pdf)
致谢: 本研究得到了四川美姑大风顶国家级自然保护区管理局的资金支持,在研究过程中得到了实验室张德军、张凯、岳先涛、陈俪心、张珮、何兴成、朱博伟、王小祎、付炎文、何梅香,贵阳市交通管理局莫名航,四川美姑大风顶国家级保护区管理局沙玛牛布、梁寅、尔石拉尔、李明、罗朝阳等人的大力支持和帮助,特此致谢;同时感谢四川大学自然博物馆郑志荣老师在鸟类分类和区划上的悉心指导!| 崔鹏, 邓文洪. 2007. 鸟类群落研究进展[J]. 动物学杂志, 42(4): 149–158. DOI:10.3969/j.issn.0250-3263.2007.04.027 |
| 戴波, 陈本平, 岳碧松, 等. 2014. 四川山鹧鸪栖息地分析与预测[J]. 四川动物, 33(3): 329–336. |
| 韩联宪, 兰道英, 马世来. 1996.云南高黎贡山鸟类多样性分布及保护[C]//中国鸟类学研究.北京: 中国林业出版社: 40-49. |
| 李操, 胡杰, 余志伟. 2003. 四川山鹧鸪的分布及生境选择[J]. 动物学杂志, 38(6): 46–51. DOI:10.3969/j.issn.0250-3263.2003.06.010 |
| 李桂垣, 张瑞云, 张清茂, 等. 1984. 四川凉山彝族自治州的鸟类区系[J]. 四川农学院学报, 2(1): 19–58. |
| 梁丹, 高歌, 王斌, 等. 2015. 云南高黎贡山中段鸟类多样性和垂直分布特征[J]. 四川动物, 34(6): 930–940. |
| 施白南, 赵尔宓. 1982. 四川资源动物志(第一卷)[M]. 成都: 四川人民出版社. |
| 斯幸峰, 丁平. 2011. 欧美陆地鸟类监测的历史、现状与我国的对策[J]. 生物多样性, 19(3): 303–310. |
| 宋昭彬, 邹方东, 郭聪, 等. 2004. 美姑大风顶自然保护区种子植物区系分析[J]. 广西植物, 24(3): 207–213. DOI:10.3969/j.issn.1000-3142.2004.03.003 |
| 吴永杰, 何兴成, DuBay SG, 等. 2017. 贡嘎山东坡的鸟类多样性和区系[J]. 四川动物, 36(6): 601–615. |
| 吴永杰, 雷富民. 2013. 物种丰富度垂直分布格局及影响机制[J]. 动物学杂志, 48(3): 797–807. |
| 张俊范. 1996. 四川鸟类鉴定手册[M]. 北京: 中国林业出版社. |
| 张荣祖. 2011. 中国动物地理[M]. 北京: 科学出版社. |
| 郑光美. 2017. 中国鸟类分类与分布名录(第三版)[M]. 北京: 科学出版社. |
| 郑作新, 潭耀匡, 罗泉笙, 等. 1962. 四川西南与云南西北地区鸟类的分类研究Ⅰ非雀形目[J]. 动物学报, 14(4): 537–554. |
| Acharya BK, Sanders NJ, Vijayan L, et al. 2011. Elevational gradients in bird diversity in the Eastern Himalaya:an evaluation of distribution patterns and their underlying mechanisms[J]. PLoS ONE, 6(12): e29097. DOI:10.1371/journal.pone.0029097 |
| Colwell RK, Mao CX, Chang J. 2004. Interpolating, extrapolating, and comparing incidence-based species accumulation curves[J]. Ecology, 85(10): 2717–2727. DOI:10.1890/03-0557 |
| MacGregor-Fors I, Schondube JE. 2011. Gray vs. green urbanization:relative importance of urban features for urban bird communities[J]. Basic and Applied Ecology, 12(4): 372–381. DOI:10.1016/j.baae.2011.04.003 |
| McCain CM. 2009. Global analysis of bird elevational diversity[J]. Global Ecology and Biogeography, 18(3): 346–360. DOI:10.1111/j.1466-8238.2008.00443.x |
| McCune B, Grace JB, Urban DL. 2002. Analysis of ecological communities[M]. Gleneden Beach, OR: MjM Software Design. |
| Mentil L, Battisti C, Maria Carpaneto G. 2018. The older the richer:significant increase in breeding bird diversity along an age gradient of different coppiced woods[J]. Web Ecology, 18(2): 143–151. |
| Pontarp M, Wiens JJ. 2017. The origin of species richness patterns along environmental gradients:uniting explanations based on time, diversification rate and carrying capacity[J]. Journal of Biogeography, 44(4): 722–735. |
| Price TD, Hooper DM, Buchanan CD, et al. 2014. Niche filling slows the diversification of Himalayan songbirds[J]. Nature, 509(7499): 222–225. DOI:10.1038/nature13272 |
| Quintero I, Jetz W. 2018. Global elevational diversity and diversification of birds[J]. Nature, 555(7695): 246–250. DOI:10.1038/nature25794 |
| Rahbek C. 2005. The role of spatial scale and the perception of large-scale species-richness patterns[J]. Ecology Letters, 8(2): 224–239. |
| Ricklefs RE. 1991. Structures and transformations of life histories[J]. Functional Ecology, 5(2): 174–183. |
| Wu YJ, Colwell RK, Rahbek C, et al. 2013. Explaining the species richness of birds along a subtropical elevational gradient in the Hengduan Mountains[J]. Journal of Biogeography, 40(12): 2310–2323. DOI:10.1111/jbi.12177 |
| Zhang Q, Hanc R, Zou F. 2011. Effects of artificial afforestation and successional stage on a lowland forest bird community in southern China[J]. Forest Ecology and Management, 261(11): 1738–1749. DOI:10.1016/j.foreco.2011.01.025 |