 2017, Vol. 36
2017, Vol. 36扩展功能
文章信息
- 陈涛, 覃事妮, 陈月华
- CHEN Tao, QIN Shini, CHEN Yuehua
- 基于鸟类功能性连接度的林地网络评价——以长沙市宁乡县中心城区为例
- The Assessment on Woodland Network Based on Functional Connectivity of Bird's Biological Characteristics:A Case Study in Ningxiang County, Changsha
- 四川动物, 2017, 36(5): 489-497
- Sichuan Journal of Zoology, 2017, 36(5): 489-497
- 10.11984/j.issn.1000-7083.20160312
-
文章历史
- 收稿日期: 2016-11-14
- 接受日期: 2017-04-10






由于城市化、工业化进程的加速发展,景观生态破碎化问题日益严峻,随之产生诸如空间异质性单一、景观格局割裂、生态调控能力逐渐减弱等一系列问题(McKinney,2008)。为了解决这些问题,区域生态规划逐渐兴起了以景观生态学原理为依托,令景观生态各要素的总量、质量、比值以及时空关系达到最优化,使景观生态中各资源组合在功能和结构上趋近最佳理想化,进而最大程度地提高区域生态的稳定性。景观连接度是当前景观生态学中着重研究的核心内容之一,其本质是研究同种或异种斑块间在生态功能和生态过程以及生态系统中的能量流、信息流与物质流的有机联系(武剑锋等,2008)。
景观功能性连接度从生物个体、种群、物种等角度出发,综合考虑某一特定生态过程的景观连接度类型,即使2个林地斑块之间并不存在实际的结构连接,但某些植物种子能利用风力从一块林地斑块传播到另一块,抑或某些飞行动物能够利用不实际存在的生态廊道,从一块林地斑块扩散至另一块,这2块生境便存在功能性连接(陈春娣等,2015)。当前城市生态格局的生境破碎化越来越严重,城市发展对原生物种的干扰也日趋严重,因此,针对特定物种利用功能性连接度对城市生态网络进行分析及整合的意义更加显著:其对解决景观空间类型破碎化和丰富景观空间异质性提供了理论基础,对优化景观空间格局与了解研究区域生态现状提供了技术指导和参考依据,对整个景观生态学进一步深化指导城市建设具有重要的现实意义(张勇等,2013)。
1 研究区域概况及指示物种选取 1.1 研究区域概况宁乡县位于湖南省长沙市西部,地处洞庭湖偏南地区,地理位置111°53′~112°46′E,27°55′~28°29′N。地势西部高东部低,南部陡北部缓,地形地貌以丘陵为主,山地、岗地和平原等为辅,沩水河贯穿其中。本次研究区域为湖南省长沙市宁乡县于《宁乡县城总体规划(2000—2020)》(2014修订)中确定面积为345.97 km2的中心城区。现状林地总面积85.72 km2,其中位于城市建设用地中的各林地总面积为12.86 km2,包括公园林地1.48 km2、生产林地0.66 km2、防护林地1.45 km2、附属林地4.28 km2、其他林地4.99 km2,具体用地类型见图 1。

|
| 图 1 研究区域土地利用现状图 Fig. 1 The map of present land-use in research region |
| |
长沙市宁乡县中心城区(城区)常见的鸟类有雀形目Passeriforme的乌鸫Turdus merula、八哥Acridotheres cristatellus、家麻雀Passer domesticus、红嘴蓝鹊Urocissa erythrorhyncha、白头鹎Pycnonotus sinensis、灰椋鸟Sturnus cineraceus,鹳形目Ciconiiformes的大白鹭Egretta alba、中白鹭Egretta intermedia、池鹭Ardeola bacchus、绿鹭Butorides striatus,鸽形目Columbiformes的山斑鸠Streptopelia orientalis等。本文通过查阅相关观测数据及生物资料,结合城区的实地调查,参照“焦点物种”理论及相关研究等(Lambeck,1997;Roberge & Angelstam,2004),确定指示物种的选取原则为:① 在人为干扰的影响下种群数量减少;② 在生态系统中的地位较为重要;③ 飞行扩散范围较广,能基本反映研究区域的景观格局;④ 考虑生态类型、种群规模、对人类干扰的耐受性等因素,所选物种能够反映大部分鸟类的特征。基于以上,本文选取灰椋鸟作为指示物种:灰椋鸟数量受人为干扰减少,其繁殖期因人类干扰造成的幼鸟死亡占47.46%(王日昕等,2002);由于其嗜吃害虫,可防止林地虫灾,也取食植物果实与种子,因此其在生态系统中的位置较为重要;另外,它对人类干扰的耐受性较低,常在各大型林地的林缘,接近农田、水塘的林缘及人为干扰较轻的公园活动,扩散范围较广。
2 研究及调查方法 2.1 鸟类数量调查方法采用样线法及样点法对研究区域中各土地覆盖类型中的典型区域进行鸟类数量调查。样线长度以足够覆盖研究区域各土地覆盖类型的典型区域为标准,调查时步行速度为1~2 km·h-1,使用双筒望远镜观察,记录样线两侧各50 m范围内灰椋鸟的数量。调查于2016年7月12日06:30—11:10,15:00—18:30进行(Wang et al., 2010)。由于鸟类在中午的活跃性较低,故此时间段不适宜调査(丁志峰等,2016)。
2.2 连接度指数评价法目前有关景观连接度研究的指数较多,如连接性指数(CI)、生态连接度指数(ECI)等,其各有不同的优缺点(邬建国,2004)。本文选取其他学者基于生境可利用性的观点提出的组分数(number of components,NC)、连接度概率指数(probability of connectivity,PC)和综合连接度指数(integral index of connectivity,IIC),利用GIS中的Conefor Sensinode 2.6插件进行数据处理与分析。其中NC用于表达功能上互相连接的斑块组成的整体数,用其来表示存在的孤立斑块的数量,值越小越好;PC和IIC通过考虑生物体的扩散距离以及各斑块面积及其所处的空间格局,可以反映出在不受人为影响的最理想情况下的景观破碎化程度(富伟等,2009),通过进一步的斑块重要性(dI)以及连线重要性(dILink)计算,直接反映斑块与连线的相对重要性,用以识别生态源地与垫脚石斑块(刘世梁等,2012),其各个指数计算公式如下:

|
(1) |
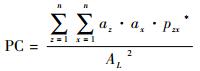
|
(2) |
式中,n为斑块的数量,az为斑块z的面积,ax为斑块x的面积,lzx为斑块z与斑块x间的连接数,pzx*为物种在斑块z与x之间扩散的最大可能性,AL为研究区域的总面积。两者计算结果的范围为:0≤IIC≤1,0<PC<1,其中,当IIC越接近1时,表明研究区域内的生态源斑块越多,越趋近于一个整体;当PC越接近1时,表明研究区域内斑块间功能性连接存在的可能性越高(姜磊等,2012)。
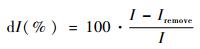
|
(3) |
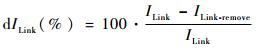
|
(4) |
式中I及ILink分别表示斑块与连线的各连接度指数,本文的连接度指数值为IIC与PC,其各自的值越高表示在该生态网络中该斑块或连线越重要。Iremove与ILink-remove分别表示将某一斑块与连线从空间中提出后的连接度指数值。
2.3 最小累积阻力评价法由于城市中鸟类的飞行扩散运动会受不同土地覆盖类型以及人为活动的干扰,现状林地的网络连接程度会低于NC、PC、IIC所得的最优化网络连接程度,而最小累积阻力模型(minimum cumulative resistance,MCR)通过考虑生态源地、扩散距离、阻碍界面,建立鸟类飞行时所受阻力的模型(Yu,1996),可以直观地描述鸟类飞行时所受的累积阻力,以此判断出林地网络当前的运行情况,计算公式如下:
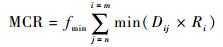
|
(5) |
式中,f为一个单调递增的函数,反映鸟类从生态源地扩散到空间中任意位置的阻力与其经过的距离和阻碍界面的正相关关系;Dij为鸟类从生态源地j飞行到位置i所用的距离;Ri为飞行所受土地覆盖类型以及人为活动的阻力。
3 研究结果 3.1 灰椋鸟数量调查结果本研究采用2条样线覆盖作为样点的7个林地斑块及2个水库以及周边的道路行道树、农田、农村建设用地及城市建设用地(图 2),观察记录沿线50 m范围内的灰椋鸟数量。其中各样地观察发现的灰椋鸟数量分别为:八家湾水库22只、林地斑块65号28只、林地斑块6号16只、林地斑块5号10只、林地斑块7号18只、白云水库16只、林地斑块62号32只、林地斑块39号8只、林地斑块28号13只,观察发现样线周边道路行道树共8只、农田及周边林缘共18只、农村建设用地共5只、城市建设用地共4只。此外以建筑物及树木为高度参照,发现灰椋鸟的最大飞行高度为20 m,通过望远镜目测的方式发现灰椋鸟的最大单次飞行距离为1 180 m。

|
| 图 2 灰椋鸟调查图 Fig. 2 Survey map of Sturnus cineraceus |
| |
本研究通过提取原有土地现状中的林地斑块导入GIS中,并选取面积>1 hm2且<5 hm2的林地斑块为小型斑块,面积>5 hm2且<10 hm2的为中型斑块,面积>10 hm2且<20 hm2的为中大型斑块,面积>20 hm2且<50 hm2的为大型斑块,面积>50 hm2的为巨型斑块(黄佳乐,2010)。然后在GIS的CS26插件中将各斑块间的连接距离设置为灰椋鸟的最大单次飞行距离1 180 m,将连接概率设定为0.5。得出城区各林地间灰椋鸟的林地网络连接图(图 3)。此时整个网络的NC值为3,即有3个独立的林地斑块组团,分别为南北大部分林地斑块,东北部由斑块58、59、60组成的组团,斑块77形成的单独林地斑块,因此整个网络没有形成完整的平面环状网络;根据公式(1)(2) 计算得出此时研究区域的IIC值与PC值分别为0.027、0.048。

|
| 图 3 林地网络图 Fig. 3 The figure of woodland network |
| |
(1) 斑块重要性分析:根据公式(3) 可得各林地斑块的重要性如图 4所示,其中,dIIC值越高表示该斑块对于整个网络的连接度贡献越大,dPC值越高表示该斑块的连接度潜力越高,越容易与周边其他林地斑块产生连接。

|
| 图 4 林地斑块重要性计算结果 Fig. 4 The importance of woodland patches |
| |
由图 4可知,共有10个林地斑块的dIIC>10,这10个林地斑块对研究区域现状的功能性连接度贡献重大,在今后的生态规划中应列为重点保护对象,最大化地减少人为因素的干扰,其中斑块43与斑块31的数值较为特别:斑块43的dIIC值为54.55,占所有dIIC值总和的22.32%,是最应该进行保护的斑块;斑块31是唯一一个dIIC>10的大型斑块,反映其位置在整个网络格局中十分重要。而1<dIIC<5的14个林地斑块同样对整个研究区域的功能性连接度有较大的贡献,应该给予一定的保护以及适当的利用。dPC值象征着各斑块与周围斑块连接的可能性,也象征着各林地的潜力,其中>5的林地斑块共13个,这些林地斑块是研究区域未来林地生态网络建设的主要依托,是研究区域踏脚石系统所围绕的核心;1<dPC<5的林地斑块共18个,这些林地斑块作为区域踏脚石系统的依托进行系统的林地系统规划。
由计算所得的dIIC值与dPC值可知,面积大小并不是dIIC值与dPC值唯一的参考依据,在整个网络格局中的位置同样对其有较大的影响;dIIC值与dPC值分别代表了林地斑块在当前林地网络中的重要性与潜力值,在今后的林地网络优化中应综合考虑这2个数值的大小,本研究根据其定义以及相关专家建议,分别赋予dIIC值0.6与dPC值0.4的权重值进行综合计算,公式为:dN=0.6dIIC+0.4dPC,计算所得的值代表了该林地斑块对于未来网络优化的重要性,根据计算结果对林地斑块进行重要性分类,将dN≥5的林地斑块作为核心林地斑块,1<dN<5的作为重要林地斑块,这些斑块起生态源作用,其余的为一般林地斑块,起连通作用(表 1)。对于核心林地斑块应作为禁建区严格控制人为因素的干扰;对于重要林地斑块应作为限建区,将人为因素的干扰尽量减少(王学海等,2013)。
| 林地斑块类型 | 斑块序号 | dN | 斑块序号 | dN |
| 核心林地斑块 | 43 | 51.98 | 49 | 11.71 |
| 50 | 28.94 | 70 | 8.72 | |
| 44 | 28.27 | 55 | 6.76 | |
| 62 | 20.06 | 65 | 5.83 | |
| 31 | 18.07 | 48 | 5.08 | |
| 52 | 13.34 | |||
| 重要林地斑块 | 42 | 4.69 | 51 | 1.76 |
| 67 | 4.53 | 72 | 1.74 | |
| 78 | 4.13 | 46 | 1.65 | |
| 64 | 3.86 | 68 | 1.49 | |
| 73 | 3.14 | 53 | 1.34 | |
| 1 | 2.73 | 54 | 1.16 | |
| 45 | 2.37 | 63 | 1.12 | |
| 61 | 1.89 |
(2) 连线重要性分析:各连线的dIICLink值反映了该连线对整个林地网络灰椋鸟飞行廊道连接度的贡献程度以及在整个生态网络的重要程度,因此可作为重要的参考依据,指导今后鸟类飞行廊道以及生态廊道的建设。根据公式(4) 将dIICLink≥1的连线作为核心飞行廊道,0.1<dIICLink<1的作为重要飞行廊道,其余的作为一般飞行廊道,其计算结果及分类见表 2。在今后的城市建设中,应该着重保护核心飞行廊道以及重要飞行廊道,并且禁止在2种飞行廊道上进行高强度的城市建设活动;当建筑高度高于鸟类平均飞行高度时应避开鸟类的飞行廊道,并保持建筑间有足够的间距,由此来降低鸟类飞行扩散时的风险(Burton et al., 2002)。
| 连线类型 | 斑块间连线 | dIICLink | 斑块间连线 | dIICLink |
| 核心飞行廊道 | 43-31 | 19.32 | 52-50 | 1.95 |
| 62-31 | 19.04 | 62-61 | 1.38 | |
| 44-43 | 3.54 | 49-43 | 1.25 | |
| 65-64 | 2.41 | 70-67 | 1.19 | |
| 49-44 | 1.99 | 67-65 | 1.19 | |
| 50-49 | 1.96 | |||
| 重要飞行廊道 | 50-42 | 0.70 | 51-50 | 0.20 |
| 78-70 | 0.60 | 78-72 | 0.15 | |
| 48-43 | 0.54 | 53-50 | 0.15 | |
| 57-55 | 0.37 | 50-45 | 0.12 | |
| 55-52 | 0.36 | 63-62 | 0.12 | |
| 56-55 | 0.35 | 49-48 | 0.11 | |
| 50-48 | 0.33 | 50-46 | 0.11 | |
| 45-43 | 0.21 | 28-3 | 0.11 |
由表 2可以发现廊道43-31与62-31的dIICLink值十分突出,结合图 3的位置显示:这2条连线是南北2个组团的重要连接廊道,对整个林地网络的物质流、能量流的贡献巨大,也从侧面反映了斑块31,即鳝鱼洲的位置在整个林地网络格局中至关重要。
3.3 灰椋鸟飞行廊道现状阻力分析鸟类飞行所受的阻力主要分为两大类型:一是土地覆盖类型。城区大部分鸟种都是林 鸟,其生活觅食环境基本相似,而土地覆盖类型与其生活环境越相似,鸟类越易进行 飞行扩散活动(Smit & Visser,1993 ;周华锋,傅伯杰,1998 ;Ortega,2012 );二是人为活动的干扰(Francis et al., 2009 )。人为活动所产生的障碍物、噪音、光污染、震动、视觉干扰等因素破坏了鸟类生 活的正常生境,且这些因素对鸟类的影响具有边缘效应,干扰的强度以干扰点为中心 向边缘递减(Hansen & Defries,2007 )。不同土地覆盖类型和人为干扰因素的阻力值由低到高排列:自然生境<半自然生 境<道路用地<建筑用地、其他道路<风景道、旅游服务中心区<市区综合服务中心 区、建筑密度<建筑高度(Lin et al., 2012 )。本研究以灰椋鸟为指示物种,以基于样线法、样点法观察所得的同等观测范围内 不同土地覆盖类型中出现的灰椋鸟数量为依据之一,通过将林地中观察所得灰椋鸟数 量作为基准,用其余土地覆盖类型中所观察的灰椋鸟数量与之进行对比,并参考其他 相关研究中人为干扰因素对鸟类的阻力系数的设立,得出综合总体的阻力系数( 表 3 ),最后通过GIS的成本距离分析工具计算得出累积阻力图,并通过自然断点法将累计 阻力图进行重分类,分为轻度阻碍、中度阻碍、重度阻碍(黎运喜等,2016 )。通过与林地网络连接图进行叠加,得出灰椋鸟的飞行阻力图(图 5)。
| 土地覆盖类型 | 数量/只 | 阻力系数 | |
| 林地 | 125 | 0 | |
| 水系及周边林缘 | 38 | 10 | |
| 农田及周边林缘 | 18 | 20 | |
| 道路行道树 | 8 | 40 | |
| 农村建设用地 | 5 | 80 | |
| 城市建设用地 | 低密度建设用地 | 2 | 100 |
| 中密度建设用地 | 2 | 130 | |
| 高密度建设用地 | 0 | 150 | |
| 建筑高度<20 m | 110 | ||
| 建筑高度>20 m | 150 | ||

|
| 图 5 灰椋鸟飞行阻力 Fig. 5 Flight resistance of Sturnus cineraceus 图中序号为林地斑块序号,红色标注的斑块序号为核心林地斑块,蓝色标注的斑块序号为重要林地斑块,黑色标注的斑块序号为一般林地斑块。 Serial number labeled in red refers to the patches of core woodland, blue refers to the patches of important woodland and black refers to the patches of common woodland. |
| |
(1) 林地斑块连接度分析:由图 5可知,核心林地斑块中大部分现状良好,少部分受不同程度的人为干扰,其中斑块65因大面积水库影响受大面积轻度干扰,但鉴于水库对其他生物的多样性具有促进作用,且丰富局部空间的异质性,故可保持现状;斑块31因周边土地覆盖类型影响及本身的面积大小受中度干扰;斑块43因居民点的人为干扰受中度干扰。重要林地斑块中,其所包含的林地斑块的受损现状对于灰椋鸟基本都属于可以接受的状况。一般林地斑块则大部分受周边土地覆盖类型以及人为干扰的影响,其生态现状呈现不同程度的受损状况,尤其位于城市建设用地中的林地斑块,受周边人为环境的干扰严重,其本身的生态现状对于灰椋鸟的吸引力极低。就整体网络而言,城区中的灰椋鸟大多只能在西北部林地斑块组团与东南部的林地斑块组团中飞行扩散。受人为因素的干扰,城区的中部与东北部的林地斑块分布处于人为阻碍孤立化与空间阻碍孤立化的状态,因此灰椋鸟不倾向于在这2个区域进行生活觅食。
(2) 飞行廊道连接度分析:结合图 5发现,大部分的核心飞行廊道与重要飞行廊道处于轻度阻碍的状态,而受城市建设用地的高密度建设用地的人为干扰,廊道50-42、28-3处于中度阻碍状态;对于一般飞行廊道,由于受城市高密度建设用地中的人为干扰影响,大部分位于城市内的林地斑块与其他林地斑块间的飞行廊道的现状较差,一共有28条处于严重阻碍的状况。
4 讨论本文通过提取宁乡县中心城区的林地斑块;并根据选取指示物种的原则选用灰椋鸟作为城区鸟类的指示物种;根据灰椋鸟的飞行特性并利用GIS的CS26插件对整个林地网络的功能性连接度指数进行计算,得出每个林地斑块与飞行廊道的连接度与重要性;根据灰椋鸟数量的分布细化各种因子的阻力系数并利用GIS的成本距离工具得出飞行累积阻力图;最后通过对各个结果的叠加分析得出主要结论,在研究的过程中,发现一些相关问题值得深入研究及探讨。
本研究通过利用连接度指数以及累积阻力模型分别确定林地网络中各斑块、连线的重要性,以及对各林地斑块、连线飞行阻力的现状分析,科学、客观地反映了各林地斑块间对于鸟类进行飞行扩散运动的连接度现状。此研究方法在充分考虑鸟类扩散现状和城市发展现状的前提下,可将城市建设活动对自然的影响最小化,从而降低了城市的无序扩张造成的自然生态环境严重恶化等问题的风险。故对于我国当前的城市林地系统格局优化以及城市建设用地边界、生态红线以及基本农田边界三线的划定具有很好的借鉴意义。
本研究通过数据分析,发现对于鸟类扩散运动而言,宁乡县中心城区的林地网络存在林地斑块及飞行廊道受不同程度人为因素的干扰,整体林地网络未形成完整的环状网络等问题,因此今后研究区域应以本研究为重要参考,通过在斑块间距大于鸟类扩散运动距离的林地斑块中建立“踏脚石”,再以新建的“踏脚石”为骨架,通过建设生物廊道等实际连接廊道构建能稳定整体生态环境、生物多样性及提高城市开放空间价值,同时提升景观整体功能的生态网络(Jongman,2003)。
本研究发现,林地网络中各林地斑块的连接度重要性并不与斑块的面积呈绝对正相关,林地斑块位于整体网络格局的位置同样重要。此外由于城市林地中鸟类的物种数并不会随林地斑块面积的增大而减少(陈水华等,2005),且主要取决于微生境的异质性(Sattler et al., 2010),故在今后城市林地系统规划中要注重林地斑块位置的选取,以及林地中的植物配置形式(杨刚等,2015)。
在对目标区域进行功能性连接度指数计算以及各因素阻力系数的设立时,对于不同的物种,往往会得出不同的林地网络连接度结果(吴昌广等,2009)。此外,距离阈值的设定与阻力系数的设定尚没有权威的方法,因此基于不同的思考方向往往会产生不一样的值,从而使整个生态网络出现不一样的计算结果,在今后的生态网络优化的工作中,应建立完整、科学、权威的优化方法论。
致谢: 感谢中南林业科技大学风景园林学院的覃事妮高级工程师、陈月华副教授对本文的指导,感谢中南林业科技大学的戴亚男、杨涛、缪鹏程、王兴月等同学协助调查灰椋鸟的数量。| 陈春娣, MeurkDC, StewartHG, 等. 2015. 城市生态网络功能性连接辨识方法[J]. 生态学报, 35(19): 6414–6424. |
| 陈水华, 丁平, 郑光美, 等. 2005. 园林鸟类群落的岛屿性格局[J]. 生态学报, 25(4): 657–663. |
| 丁志锋, 梁健超, 陈本亮, 等. 2016. 广州市园林鸟类物种和功能多样性的变化[J]. 四川动物, 35(5): 759–764. |
| 富伟, 刘世梁, 崔保山, 等. 2009. 景观生态学中生态连接度研究进展[J]. 生态学报, 29(11): 6174–6182. DOI:10.3321/j.issn:1000-0933.2009.11.052 |
| 黄佳乐. 2010. 重庆市主城区森林斑块特征分析与格局优化[D]. 重庆: 西南大学. http://cdmd.cnki.com.cn/Article/CDMD-10635-2010094421.htm |
| 姜磊, 岳德鹏, 曹睿, 等. 2012. 北京市朝阳区景观连接度距离阈值研究[J]. 林业调查规划, 2: 18–22, 32. DOI:10.3969/j.issn.1671-3168.2012.01.005 |
| 黎运喜, 陈佑平, 余凌帆, 等. 2016. 基于MaxEnt模型的二郎山廊道大熊猫栖息地适宜性评价[J]. 四川动物, 35(6): 833–837. |
| 刘世梁, 杨珏婕, 安晨, 等. 2012. 基于景观连接度的土地整理生态效应评价[J]. 生态学杂志, 31(3): 689–695. |
| 王日昕, 车轶, 石戈, 等. 2002. 灰椋鸟的繁殖生态[J]. 动物学杂志, 1: 58–59. DOI:10.3969/j.issn.0250-3263.2002.01.015 |
| 王学海, 和艳, 李迎彬. 2013. 城市禁建区规划实践——以昆明中心城区禁建区划定为例[J]. 城市规划, 37(4): 43–49. |
| 邬建国. 2004. 景观生态学中的十大研究论题[J]. 生态学报, 24(9): 2074–2076. |
| 吴昌广, 周志翔, 王鹏程, 等. 2009. 基于最小费用模型的景观连接度评价[J]. 应用生态学报, 20(8): 2042–2048. |
| 武剑锋, 曾辉, 刘雅琴. 2008. 深圳地区景观生态连接度评估[J]. 生态学报, 28(4): 1691–1701. |
| 杨刚, 王勇, 许洁, 等. 2015. 城市公园生境类型对鸟类群落的影响[J]. 生态学报, 35(12): 4186–4195. |
| 张勇, 屠宁雯, 姚林泉. 2013. 城市道路交通网络脆弱性辨识方法[J]. 中国公路学报, 26(4): 154–161. |
| 周华锋, 傅伯杰. 1998. 景观生态结构与生物多样性保护[J]. 地理科学, 18(5): 472–479. |
| Burton NHK, Armitage MJS, Musgrove AJ, et al. 2002. Impacts of man-made landscape features on numbers of estuarine water birds at low tide[J]. Environmental Management, 30: 857–864. DOI:10.1007/s00267-002-2732-5 |
| Francis CD, Ortega CP, Cruz A. 2009. Noise pollution changes avian communities and species interactions[J]. Current Biology, 19: 1415–1419. DOI:10.1016/j.cub.2009.06.052 |
| Hansen AJ, Defries R. 2007. Ecological mechanisms linking protected areas to surrounding lands[J]. Ecological Applications, 17: 974–988. DOI:10.1890/05-1098 |
| Jongman RHG. 2003. Ecological networks and greenways in Europe:reasoning and concepts[J]. Journal of Environmental Sciences, 15(2): 173–181. |
| Lambeck RJ. 1997. Focal species:a multi-species umbrella for nature conservation[J]. Conservation Biology, 11: 849–856. DOI:10.1046/j.1523-1739.1997.96319.x |
| Lin T, Coppack T, Lin QX, et al. 2012. Does avian flight initiation distance indicate tolerance towards urban disturbance[J]. Ecological Indicators, 15: 30–35. DOI:10.1016/j.ecolind.2011.09.018 |
| McKinney ML. 2008. Effects of urbanization on species richness:a review of plants and animals[J]. Urban Ecosystems, 11(2): 161–176. DOI:10.1007/s11252-007-0045-4 |
| Ortega CP. 2012. Effects of noise pollution on birds:a brief review of our knowledge[J]. Ornithological Monographs, 74: 6–22. DOI:10.1525/om.2012.74.1.6 |
| Roberge JM, Angelstam P. 2004. Usefulness of the umbrella species concept as a conservation tool[J]. Conservation Biology, 18: 76–85. DOI:10.1111/cbi.2004.18.issue-1 |
| Sattler T, Borcard D, Arlettaz R, et al. 2010. Spider, bee, and bird communities in cities are shaped by environmental control and high stochasticity[J]. Ecology, 91(11): 3343–3353. DOI:10.1890/09-1810.1 |
| Smit CJ, Visser GJM. 1993. Effects of disturbance on shorebirds:a summary of existing knowledge from the Dutch Wadden Sea and Delta area[J]. Wader Study Group Bulletin, 68: 6–19. |
| Wang YP, Bao YX, Yu MJ, et al. 2010. Nestedness for different reasons:the distributions of birds, lizards and small mammals on islands of an inundated lake[J]. Diversity and Distributions, 16(5): 862–873. DOI:10.1111/j.1472-4642.2010.00682.x |
| Yu KJ. 1996. Security patterns and surface model in landscape ecological planning[J]. Landscape and Urban Planning, 36: 1–17. DOI:10.1016/S0169-2046(96)00331-3 |