 2017, Vol. 36
2017, Vol. 36扩展功能
文章信息
- 张源笙, 蒋健, 蒋万杰, 王丹, 范雅倩, 汤小明, 鲍伟东
- ZHANG Yuansheng, JIANG Jian, JIANG Wanjie, WANG Dan, FAN Yaqian, TANG Xiaoming, BAO Weidong
- 北京松山国家级自然保护区兽类活动节律初步研究
- Activity Patterns of Mammals in Beijing Songshan National Nature Reserve
- 四川动物, 2017, 36(4): 460-467
- Sichuan Journal of Zoology, 2017, 36(4): 460-467
- 10.11984/j.issn.1000-7083.20170108
-
文章历史
- 收稿日期: 2017-04-04
- 接受日期: 2017-05-23






2. 北京市松山国家级自然保护区管理处, 北京 102115
2. Songshan National Nature Reserve Management Department, Beijing 102115, China
作为动物行为生态学研究的重要内容,活动节律的研究主要是通过探查动物活动强度的时空变化及其规律的影响因素等来了解动物的生存行为策略(尚玉昌,2006)。野生动物的行为模式具有明显的季节性差异和昼夜节律(Nielsen,1984),活动节律既是动物自身长期进化的结果(张世卿,1995),又体现了对环境因素变化的适应能力(Risenhoover,1986;Shi et al., 2003),其受生物学因素(如年龄、性别、繁殖状态等)和外界环境因素(如气候、光照、捕食者、竞争者、食物资源等)的共同作用,与环境周期变化保持同步(尚玉昌,2006;段利娟,2014)。因此,开展动物行为生态学研究、分析动物的生存策略,需要首先了解动物的行为节律和活动时间分配(胡亮,2015)。
北京松山国家级自然保护区曾分别于2010年和2013年开展过2次红外相机野生动物资源多样性调查,已对现有物种构成有基本了解(刘芳等,2012),然而尚未在此基础上进行更深入的生态学研究。本文利用2013年3月—2014年2月的红外相机监测数据,探究保护区内10种最常见兽类的活动节律,为制定区域性生物多样性保护措施提供数据支持,并分析红外相机可能对兽类行为产生的干扰效应,探讨这些行为对使用红外相机监测野生动物中存在的问题和解决方案。
1 研究方法 1.1 研究地区北京松山国家级自然保护区位于北京市延庆县西北部大海坨山南麓,115°43′44″~115°50′22″E,40°29′9″~40°33′35″N,总面积4 671 hm2(刘芳等,2012)。保护区地处切割强烈的中山地带,海拔627~2 198 m,地势起伏较大。全区年最高气温39 ℃,最低气温-27.3 ℃,年均气温8.5 ℃,年均降水量493 mm,年蒸发量1 772 mm,森林覆盖率87.7%(齐磊等,2012)。由于地形差异形成小气候的多样性和复杂性,生物多样性丰富(崔海鸥,单宏臣,2006),是华北地区保存较好的暖温带山地森林生态系统和野生动植物类型自然保护区(张晓秋,2004;齐磊等,2012)。保护区拥有华北地区唯一的天然油松林,面积达50 hm2(高杰等,2016)。动物区系组成以古北界物种为主,记载有脊椎动物68科216种及亚种,其中兽类15科29种,鸟类37科119种(杜连海等,2012)。
1.2 红外相机布设方法以提高动物拍摄率为目标,根据2010年调查的动物分布信息,以及现场实地发现的粪便、足迹、遗落物、取食和挖掘痕迹等,将红外相机布设在兽径、水源点附近等动物活动较多且视野开阔的区域(图 1)。相机固定于离地面50~80 cm的树干上,镜头与地面呈小于10°的俯角,以扩大拍摄视野,同时有效防止正午时地面反射光线过强,过度曝光触发相机连续拍摄。

|
| 图 1 北京松山国家级自然保护区红外相机布设位点 Fig. 1 The location of infrared camera sites in Beijing Songshan National Nature Reserve |
| |
本研究使用64台Ltl Acorn 6210数码红外相机,累计布放位点384个。拍照像素为500万,视频尺寸为1080P,拍摄模式为连拍3张,再录制一段10~60 s视频,2次启动间隔3 s,24 h监测。布设相机时记录放置的时间、经纬度、海拔、坡度、坡向、植被类型、动物痕迹、人为干扰强度等信息。每月更换电池并导出数据,再将红外相机移至其他位点。
1.3 数据处理方法根据松山的气候特征,四季划分为:春季3—5月、夏季6—8月、秋季9—11月、冬季12月—翌年2月(杜连海等,2012)。以独立事件和相对丰富度(relative abundance index,RAI)指数作为动物活动节律的指标(武鹏峰等,2012)。依据红外相机拍摄影像,本文从中选取相对丰富度最高的10种兽类为研究对象,独立事件共计2 247组。
季节活动节律,以各月相对丰富度(MRAI)为指标,采用公式计算: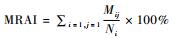
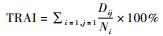
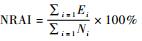
数据在Excel 2013和SSPS 20.0中处理分析。某物种相对丰富度的季节性差异用独立样本的Kruskal-Wallis H检验。显著性水平设置为α=0.05。
2 研究结果 2.1 季节活动节律结果表明,季节因素对岩松鼠Sciurotamias davidianus(Z=54.337,n=3,P<0.001)、亚洲狗獾Meles leucurus(Z=72.213,n=3,P<0.001)、猪獾Arctonyx collaris(Z=65.898,n=3,P<0.001)、豹猫Prionailurus bengalensis(Z=37.787,n=3,P<0.001)、貉Nyctereutes procyonoides(Z=24.344,n=3,P<0.001)、北花松鼠Tamias sibiricus(Z=61.851,n=3,P<0.001)、中华斑羚Naemorhedus griseus(Z=25.070,n=3,P<0.001) 和野猪Sus scrofa(Z=8.377,n=3,P=0.039) 的独立事件数影响显著,而对果子狸Paguma larvata(Z=3.868,n=3,P=0.276) 影响不显著。
利用月相对丰富度分析9种兽类的季节活动节律(表 1)。在小型兽类中,北花松鼠和岩松鼠的活动节律相似,在夏末至秋季活动频繁,活动高峰集中于7月和9月。在5种中型兽类中,猪獾和亚洲狗獾的活动节律相似,夏季、秋季相对丰富度较高,冬季则相对较低,活动高峰分别集中在8月和11月;貉和豹猫的活动节律相似,夏季、初秋相对丰富度较高,其他时间分布均匀,活动高峰分别集中在7月和8月;而果子狸秋季相对丰富度较高,冬季则相对较低,活动高峰集中在11月。在2种大型兽类中,中华斑羚和野猪活动节律相似,秋季相对丰富度较高,夏季则相对较低,活动高峰都集中在11月,野猪在3月还有1个活动小高峰。
| 物种 Species | 独立事件数 RAI | 2013年 | 2014年 | ||||||||||
| 3月 | 4月 | 5月 | 6月 | 7月 | 8月 | 9月 | 10月 | 11月 | 12月 | 1月 | 2月 | ||
| 岩松鼠 Sciurotamias davidianus | 1 104 | 2.90 | 2.36 | 1.81 | 3.17 | 4.17 | 21.56 | 40.76 | 11.05 | 5.62 | 1.72 | 2.17 | 2.72 |
| 亚洲狗獾 Meles leucurus | 269 | 0.74 | 0 | 16.73 | 10.78 | 14.13 | 18.96 | 9.29 | 0.37 | 27.88 | 0.37 | 0 | 0.74 |
| 猪獾 Arctonyx collaris | 247 | 3.64 | 4.45 | 15.38 | 11.34 | 11.74 | 26.32 | 12.96 | 11.74 | 1.21 | 0.40 | 0 | 0.81 |
| 豹猫 Prionailurus bengalensis | 235 | 2.13 | 2.13 | 2.13 | 4.26 | 10.21 | 47.23 | 9.36 | 6.38 | 4.26 | 3.83 | 2.13 | 5.96 |
| 貉 Nyctereutes procyonoides | 112 | 5.36 | 0.89 | 2.68 | 5.36 | 33.04 | 21.43 | 9.82 | 4.46 | 9.82 | 1.79 | 2.68 | 2.68 |
| 北花松鼠 Tamias sibiricus | 107 | 3.74 | 0 | 0 | 6.54 | 27.10 | 24.30 | 21.50 | 16.82 | 0 | 0 | 0 | 0 |
| 果子狸 Paguma larvata | 52 | 0 | 3.85 | 3.85 | 1.92 | 3.85 | 1.92 | 0 | 1.92 | 82.69 | 0 | 0 | 0 |
| 中华斑羚 Naemorhedus griseus | 33 | 0 | 3.03 | 0 | 0 | 0 | 0 | 0 | 15.15 | 54.55 | 27.27 | 0 | 0 |
| 野猪 Sus scrofa | 32 | 34.38 | 0 | 0 | 0 | 0 | 6.25 | 0 | 3.13 | 34.38 | 3.13 | 6.25 | 12.50 |
从全年数据分析,北花松鼠和岩松鼠的活动主要集中在昼间,日活动曲线均呈单峰型。其中,北花松鼠的活动高峰集中在08: 00—14: 00,峰值出现在10: 00—12: 00;岩松鼠的活动高峰集中在08: 00—14: 00,峰值出现在08: 00—10: 00。从秋末到冬季的数据分析,松鼠Sciurus vulgaris的活动高峰集中在06: 00—12: 00,峰值出现在08: 00—10: 00(图 2)。

|
| 图 2 北花松鼠、松鼠、岩松鼠的日活动节律 Fig. 2 The daily activity patterns of Tamias sibiricus, Sciurus vulgaris, and Sciurotamias davidianus |
| |
对岩松鼠在秋、冬季的活动情况分析发现,秋季在昼间均有较强活动,持续时间为07: 00—17: 00;但在冬季则出现几个活动高峰,除在08: 00—15: 00持续活动外,在06: 00—07: 00和16: 00—17: 00有2个小高峰(图 3)。另外,秋季和冬季的最大活动峰值均出现在09: 00—10: 00。

|
| 图 3 岩松鼠秋季和冬季的日活动节律 Fig. 3 The daily activity patterns of Sciurotamias davidianus in autumn and winter |
| |
豹猫、亚洲狗獾、猪獾和貉夜间活动频繁。豹猫的日活动曲线呈双峰型,活动高峰集中在18: 00—24: 00,在02: 00—04: 00有1个小高峰。狗獾和猪獾的日活动曲线呈单峰型,活动高峰分别集中在16: 00—22: 00和20: 00—04: 00,其他时间活动频率较平均。貉的日活动曲线呈三峰型,活动高峰集中在18: 00—20: 00,在04: 00—06: 00、14: 00—16: 00有2个小高峰(图 4)。

|
| 图 4 豹猫、亚洲狗獾、猪獾和貉的日活动节律 Fig. 4 The daily activity patterns of Prionailurus bengalensis, Meles leucurus, Arctonyx collaris, and Nyctereutes procyonoides |
| |
果子狸、野猪和中华斑羚的日活动节律多集中在晨昏和夜间。果子狸的日活动曲线呈单峰型,活动高峰集中在20: 00—06: 00。野猪的日活动曲线呈双峰型,活动高峰集中在16: 00—22: 00,在04: 00—06: 00有1个小高峰。中华斑羚的日活动曲线呈双峰型,活动高峰集中在20: 00—04: 00,在06: 00—08: 00有1个小高峰(图 5)。

|
| 图 5 果子狸、野猪和中华斑羚的日活动节律 Fig. 5 The daily activity patterns of Paguma larvata, Sus scrofa, and Naemorhedus griseus |
| |
利用夜间相对丰富度分析10种兽类的夜行性特点,发现果子狸、中华斑羚、猪獾、豹猫和野猪的夜间相对丰富度明显高于13/24,说明夜间活动能力较强,属于夜行性。貉和狗獾的夜间相对丰富度接近13/24,夜行性不明显。松鼠、北花松鼠和岩松鼠的夜间相对丰富度明显低于13/24,说明昼间活动能力较强,具有昼行性(图 6)。

|
| 图 6 北京松山国家级自然保护区10种常见兽类的夜间相对丰富度 Fig. 6 The NRAI of the 10 most-common mammal species in Beijing Songshan National Nature Reserve 虚线对应的纵坐标为13/24(约54.20%)。 The ordinate value of vertical line is 13/24 (about 54.20%). |
| |
调查中记录到兽类伫立观望、警戒、凑近、嗅闻、舔舐、探查红外相机等行为的独立事件共计48组。按兽类发现红外相机的独立事件数占该物种独立事件数的比例由高到低依次为:中华斑羚(12.12%)、野猪(9.38%)、豹猫(6.38%)、貉(6.25%)、狗獾(4.83%)、猪獾(2.43%),显示大型动物对红外相机较为敏感。
3 讨论在森林生态系统中,植被茂密限制视野,以及部分野生动物生性警觉、行踪隐蔽、昼伏夜出等,用传统调查法研究动物活动节律困难。红外相机技术具有全时性、标准化、非损伤、抗环境变化,易获得大型兽类、夜行性及高隐蔽性物种信息的能力(O'Connell et al., 2011),且安全、人力成本低等诸多优势,在研究林地兽类动物活动节律中发挥了重要作用(肖治术等,2014;张履冰等,2014)。
类活动节律的季节性差异进行研究,结果发现,所调查的9种兽类在研究区域内相对丰富度达95.51%。岩松鼠和北花松鼠的独立事件数之和占兽类独立事件数的52.80%,岩松鼠的相对丰富度(33.53%)明显高于其他兽类,是本地区的优势物种。保护区内的大片天然原始油松林为松鼠科Sciuridae动物提供了大量的优质食物。
本研究结果表明,保护区内兽类的相对丰富度存在季节性差异,推测与不同物种所需食物资源的季节分布和繁殖期有关。岩松鼠和北花松鼠夏、秋季活动较为频繁,与其贮食种子的习性有关:冬、春季该区域的温度较低,过多的巢外活动会消耗大量能量,所以此时活动强度远低于夏、秋季,而选择半冬眠的方式在巢内度过寒冷时期,是松鼠科物种长期进化的适应策略(任娟等,2010;李俊生等,2013)。此外,本研究还记录到保护区此前未发现的新物种——松鼠(陈卫,高武,1991;杜连海等,2012;刘芳等,2012)。松鼠在2013年3—10月未记录到,2013年11月开始有记录,月相对丰富度依次为:2013年11月(46.43%),12月(19.64%),2014年1月(3.57%),2月(30.35%)。拍摄到松鼠的相机位置从2013年11月的3个位点逐渐扩大到2014年2月的10个位点,推测松鼠为人为放生到保护区的新物种。在保护区顺利度过第一个冬季后,松鼠是否会对原有近缘种产生入侵竞争,尚需进一步关注。松鼠科的3个物种在栖息地利用和食性选择上具有相似性(路纪琪,张知彬,2005;李俊生等,2013;杨慧,2013),根据生态位的重叠随着体型差距的减小而增大的理论(Woodward et al., 2002),三者之间可能存在高度的种间竞争。
狗獾、猪獾、豹猫、貉在夏季活动较为频繁,源于这4种兽类均为杂食动物(鲍伟东等,2005)。在夏季,植物性和动物性食物都较为丰富,采用增加活动频率、大量觅食的策略,可为越冬储存足够的脂肪。果子狸虽然也是杂食性动物,但以野果和植物种籽为主要食物(王健等,2008),因此在秋季植物结果时活动较为频繁。
中华斑羚和野猪在秋季活动较为频繁,一是为过冬摄取更多能量,二是秋末冬初正值中华斑羚的繁殖期,增加活动频率可以提高异性遇见率,说明活动节律还与繁殖期密切相关。
日活动节律和夜行性研究表明,松山保护区兽类在各时间段的相对丰富度不同,推测是由于哺乳动物内源性的昼夜节律对环境的高度适应(安扬,徐璎,2015)。松鼠科的3个物种都是典型的昼行性动物,以往研究发现影响该类动物活动节律的主要影响因子是光周期和温度变化(Tonkin,1983;Wauters et al., 1992)。松鼠在秋季的日活动时间明显多于日照时间相对较短、气温较低的冬季(李俊生等,2013)。本研究发现岩松鼠秋季日活动时间与冬季相同,只是冬季第一活动高峰的起始和结束时间比秋季更向正午集中,从而利用最适宜的温度条件。亚洲狗獾过去被认为是典型的夜行性动物(高耀亭等,1987),上海地区的研究发现,在秋季亚洲狗獾最早出洞时间可提前至16: 50,可能与其秋季食物需求量大有关(崔勇勇,2013)。本研究中亚洲狗獾的活动高峰在16: 00—22: 00,可能是为了减少与猪獾(活动高峰22: 00—04: 00) 的种间竞争。果子狸的活动高峰集中在20: 00—06: 00,是典型的夜行性动物。多次记录到2~4只果子狸排队经过相机的场面,带头个体伫立嗅闻,并用腺体涂擦地面做标记,这与陕西地区野生果子狸的集群活动和领域标记行为相符(张保良,1997)。秦岭地区的中华斑羚和陕西观音山地区的鬣羚Capricornis sumatraensis等有蹄类动物都具有明显的晨昏活动习性(武鹏峰等,2012;贾晓东等,2014)。但本研究发现中华斑羚的2个活动高峰分别在06: 00—08: 00和20: 00—04: 00,且第二个活动高峰持续时间长于第一个,峰值也更高。中华斑羚采取夜间活动的策略可能是为避开人类活动,同时也说明其具有一定的夜行能力。
本研究表明,中华斑羚、野猪、豹猫和貉等夜行性动物发现相机的比例较高。可能是因为这些动物夜视能力强(Newbold & King,2009)、听觉或嗅觉较为灵敏(Meek et al., 2014),能够发现相机发射的红外线或者散发的塑料或金属气味,以及留在红外相机上人类的气味(Séquin et al., 2003;Larrucea et al., 2007)。检查影像资料时发现,中华斑羚在经过红外相机时,有注视并靠近红外相机以及前后转动耳朵探听的行为,推测其夜视能力和听力较为发达。野猪在探测到相机的红外光后会伫立凝视红外相机2~3 s,随后快速逃离。红外相机产生的这些潜在影响因素或使动物警戒避开,或将其诱引靠近,都会导致拍摄率发生变化。此外,本研究还发现狗獾和猪獾在觅食过程中能够发现红外相机,并会直立上身用前肢和头部嗅闻红外相机并进行接触性探查。此前对新疆阿勒泰地区的马鹿Cervus elaphus和四川唐家河地区的大熊猫Ailuropoda melanoleuca的研究也发现动物由于好奇玩耍,甚至弄坏相机(张履冰等,2014)。可见,红外相机的出现会引起部分动物的探究行为。因此,虽然红外相机对动物干扰小(O'Connell et al., 2011),但仍存在影响动物拍摄率和行为的问题,使自然状态下的数据采集产生一定程度的偏差,在利用红外相机研究野生动物行为时需要考虑此类影响因素
致谢: 感谢北京松山国家级自然保护区野生动物巡护队对本研究的大力支持与配合。| 安扬, 徐璎. 2015. 哺乳动物昼夜节律机制研究进展[J]. 生命科学, 27(11): 1372–1379. |
| 鲍伟东, 李晓京, 史阳. 2005. 北京市三个区域食肉类动物食性的比较分析[J]. 动物学研究, 26(2): 118–122. |
| 陈卫, 高武. 1991. 北京松山自然保护区兽类调查报告[J]. 北京师范学院学报(自然科学版), 12(1): 64–69. |
| 崔海鸥, 单宏臣. 2006. 北京松山国家级自然保护区脊椎动物区系初报[J]. 四川动物, 25(4): 776–778. |
| 崔勇勇. 2013. 引入狗獾(Meles meles)活动规律及行为的研究[D]. 上海: 华东师范大学: 22-23. http://cdmd.cnki.com.cn/Article/CDMD-10269-1013272605.htm |
| 杜连海, 王小平, 陈峻崎, 等. 2012. 北京松山自然保护区综合科学考察报告[M]. 北京: 中国林业出版社: 1-10, 105. |
| 段利娟. 2014. 王朗自然保护区大熊猫及其同域物种活动节律及栖息地利用研究[D]. 北京: 北京林业大学: 6-7. http://cdmd.cnki.com.cn/article/cdmd-10022-1014319082.htm |
| 高杰, 郭子健, 刘艳红. 2016. 北京松山不同龄级天然油松林生物量分配格局及其影响因子[J]. 生态学杂志, 35(6): 1475–1480. |
| 高耀亭, 等. 1987. 中国动物志兽纲第八卷食肉目[M]. 北京: 科学出版: 214-223. |
| 胡亮. 2015. 基于红外相机技术的北山羊(Capra ibex)活动节律与集群行为的研究[D]. 新疆: 石河子大学: 2-3. http://cdmd.cnki.com.cn/Article/CDMD-10759-1016704930.htm |
| 贾晓东, 刘雪华, 杨兴中, 等. 2014. 利用红外相机技术分析秦岭有蹄类动物活动节律的季节性差异[J]. 生物多样性, 22(6): 737–745. |
| 李俊生, 马建章, 宋延龄. 2003. 松鼠秋冬季节日活动节律的初步研究[J]. 动物学杂志, 38(1): 33–37. |
| 刘芳, 李迪强, 吴记贵, 等. 2012. 利用红外相机调查北京松山国家级自然保护区的野生动物物种[J]. 生态学报, 32(3): 730–739. |
| 路纪琪, 张知彬. 2005. 岩松鼠的食物贮藏行为(英文)[J]. 动物学报, 51(3): 376–382. |
| 齐磊, 胡德夫, 丁长青, 等. 2012. 北京松山国家级自然保护区鼠类群落多样性与结构变动分析[J]. 林业科学, 48(9): 181–185. DOI:10.11707/j.1001-7488.20120928 |
| 任娟, 曹晓莉, 宋鹏飞, 等. 2010. 人工林赤腹松鼠春夏季活动节律与行为特征观察[J]. 四川动物, 29(6): 862–867. |
| 尚玉昌. 2006. 动物的行为节律[J]. 生物学通报, 41(10): 8–10. DOI:10.3969/j.issn.0006-3193.2006.10.004 |
| 王健, 刘群秀, 索建中, 等. 2008. 湖北后河自然保护区果子狸食物组成初步研究[J]. 动物学杂志, 43(2): 91–95. |
| 武鹏峰, 刘雪华, 蔡琼, 等. 2012. 红外相机技术在陕西观音山自然保护区兽类监测研究中的应用[J]. 兽类学报, 32(1): 67–71. |
| 肖治术, 李欣海, 王学志, 等. 2014. 探讨我国森林野生动物红外相机监测规范[J]. 生物多样性, 22(6): 704–711. |
| 杨慧. 2013. 阔叶红松林中花鼠的种群动态及生境选择特征[D]. 黑龙江: 东北林业大学: 1-31. http://cdmd.cnki.com.cn/Article/CDMD-10225-1013366720.htm |
| 张保良. 1997. 花面狸生态习性的观察[J]. 经济动物学报, 1(4): 36–41. |
| 张履冰, 崔绍朋, 黄元骏, 等. 2014. 红外相机技术在我国野生动物监测中的应用:问题与限制[J]. 生物多样性, 22(6): 696–703. |
| 张世卿. 1995. 动物的节律性活动[J]. 天中学刊, 10(1): 49–53. |
| 张晓秋. 2004. 松山自然保护区生物多样性使用价值评估[D]. 北京: 中国林业科学研究院. http://cdmd.cnki.com.cn/Article/CDMD-82201-2005030736.htm |
| Larrucea ES, Brussard PF, Jaeger MM, et al. 2007. Cameras, coyotes and the assumption of equal detectability[J]. Journal of Wildlife Management, 71(5): 1682–1689. DOI:10.2193/2006-407 |
| Meek PD, Ballard GA, Fleming PJS, et al. 2014. Camera traps can be heard and seen by animals[J]. PLoS ONE, 9(10). DOI:10.1371/jornal.pone.0110832 |
| Newbold HG, King CM. 2009. Can a predator see'invisible'light? Infrared vision in ferrets (Mustela furo)[J]. Wildlife Research, 36(4): 309–318. DOI:10.1071/WR08083 |
| Nielsen ET. 1984. Relation of behavioural activity rhythms to the changes of day and night. A revision of views[J]. Behaviour, 89(1/2): 147–173. |
| O'Connell AF, Nichols JD, Karanth KU. 2011. Camera traps in animal ecology:methods and analyses[M]. New York: Springer: 1-263. |
| Risenhoover KL. 1986. Winter activity patterns of moose in interior Alaska[J]. Wildlife Management, 50(4): 727–734. DOI:10.2307/3800990 |
| Séquin ES, Jarger MM, Brussard PF, et al. 2003. Wariness of coyotes to camera traps relative to social status and territory boundaries[J]. Canadian Journal of Zoology, 81(12): 2015–2025. DOI:10.1139/z03-204 |
| Shi J, Dunbar RIM, Buckland D, et al. 2003. Daytime activity budgets of feral goats (Capra hircus) on the Isle of Rum:influence of season, age, and sex[J]. Canadian Journal of Zoology, 81(5): 803–815. DOI:10.1139/z03-055 |
| Tonkin JM. 1983. Activity patterns of the red squirrel (Sciurus vulgaris)[J]. Mammal Review, 13(2-4): 99–111. DOI:10.1111/j.1365-2907.1983.tb00271.x |
| Wauters L, Cswinnen C, Dhondt AA. 1992. Activity budget and foraging behavior of red squirrels (Sciurus vulgaris) in coniferous and deciduous habitats[J]. Journal of Zoology, 227(1): 71–86. DOI:10.1111/jzo.1992.227.issue-1 |
| Woodward G, Jones J, Hildrew AG. 2002. Community persistence in Broadstone Stream (U.K.) over three decades[J]. Freshwater Biology, 47(8): 1419–1435. DOI:10.1046/j.1365-2427.2002.00872.x |