 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 廖阳, 闫荣玲, 陈滐, 罗金强, 李金亮
- LIAO Yang, YAN Rongling, CHEN Jie, LUO Jinqiang, Li Jinliang
- 潇水河永州零陵段沿岸蝙蝠物种多样性调查
- Preliminary Survey on Bat Diversity along Xiaoshui River Coast in Lingling Section of Yongzhou, Hunan Province
- 四川动物, 2015, 34: 370-374
- Sichuan Journal of Zoology, 2015, 34: 370-374
- 10.3969/j.issn.1000-7083.2015.03.008
-
文章历史
- 收稿日期:2014-07-08
- 接受日期:2014-11-17






蝙蝠呈世界性分布,是进化中前肢演化为翼、唯一具有飞行本领的哺乳动物类群(Altringham,1996)。蝙蝠种类繁多,目前全世界已发现的种类超千种,10年前我国已经记录120种,分属7科29属(王应祥,2003;Wilson & Reeder,2005)。近年来,国内多个研究团队开展了蝙蝠资源调查(黎道洪,罗泰昌,2002;吴毅等,2002;岑业文,彭红元,2010;黄继展等,2013)。高鞍菊头蝠Rhinolophus paradoxolophus、小褐菊头蝠R. stheno、马氏菊头蝠R. marshalli、小蹄蝠Hipposideros cineraceus、小巨足蝠Myotis hasseltii、大趾鼠耳蝠M. macrodacrtylus、马来假吸血蝠Megaderma spasma等近10个我国蝙蝠种类新纪录相继被报道(赵辉华等,2002;吴毅等,2004;张礼标等,2004,2010;张劲硕等,2005;江廷磊等,2008;谭敏等,2009)。另有多个蝙蝠新种在我国被发现、鉴定与命名,如北京宽耳蝠Barbastella beijingensis、华南菊头蝠R. huananus、西南菊头蝠R. xinanzhongguoensis、施氏菊头蝠R. schnitzleris等(Zhang et al,2007;Wu et al,2008;Zhou et al,2009;Wu & Thong,2011)。这些工作进一步丰富了我国蝙蝠种类的多样性,增加了人们对我国蝙蝠资源现状与地理分布的了解。
由于蝙蝠捕食大量农业害虫(大部分蝙蝠以昆虫为食),且对植物授粉和种子传播(部分蝙蝠如狐蝠、犬蝠等以植物花蜜和果实为食)有积极作用,因此在农林生态系统中扮演着举足轻重的角色(韩宝银,贺红早,2012)。蝙蝠种类及群体数量的多寡一定程度上反映了当地生态环境的优劣。永州位于湖南省西南部,是湘、粤、桂三省结合部,属亚热带大陆性季风气候,四季分明,辖区内生态环境良好,适合蝙蝠栖息繁衍,存在一定规模的蝙蝠种群。近年来,在永州发现和报道了多个湖南省蝙蝠种类新纪录(罗丽等,2011;李艳丽等,2012),但未有专门针对永州某一区域蝙蝠资源调查的报道,也鲜见对同一河流沿岸不同区段所栖蝙蝠种类比较的报道。本文在长期调查基础上,对潇水河永州零陵段蝙蝠物种多样性进行了初步调查,比较了上、下游不同区段蝙蝠种类的差异并分析其与生境的关系。 1 研究地点与方法 1.1 研究地点
湖南省永州市零陵区(26°12'N,111°36'E)位于湘江上游,气候温和、雨量充沛,全年平均气温在17.6~18.6 ℃之间,无霜期年均285~311 d,年降雨量1290~1900 mm。潇水是湘江上游最大的支流,其最后一段约25 km流经零陵,且在零陵城郊与湘江汇合,"潇湘"由此而来。潇水零陵段平均河宽约300 m,两岸低山起伏伴有岩壁显露,植被茂盛且四季常青,另有民居与小面积农田或菜地散布其中。下游最末端约10 km进入城郊,各类建筑物明显增多,但河西沿岸有朝阳公园、朝阳岩、柳子庙、怀素公园等景观,另有湖南科技学院、永州市第七中学及湖南中烟工业有限责任公司零陵卷烟厂等绿化率较高、污染与噪声较少的企事业单位紧邻河畔。 1.2 调查方法
2011年5月—2013年8月,对潇水零陵段沿岸蝙蝠资源进行了调查。前期对潇水河零陵段沿岸进行走访调查,了解沿岸生态及蝙蝠栖息地与活动聚集地并作好记录,其中聚集地是在傍晚时分通过肉眼和望远镜对沿岸进行观察来确定(廖阳等,2014)。调查走访结束后,对蝙蝠栖息地与活动聚集地进行样本捕捉、种类鉴定及数量统计。由于不同种类蝙蝠栖息环境及活动场所的差异,所采取的调查和样本捕捉的方法不同。小型洞穴与岩壁等通过手电照射观察并于傍晚时分在出口进行计数和张网捕捉,洞口较大者则直接进入洞穴进行捕捉和计数;栖息于竹丛与树冠的蝙蝠直接通过肉眼或借助望远镜观察与计数,并在竹丛与树冠四周布设雾网捕捉;藏于建筑物内的蝙蝠则在白天通过肉眼观察计数并通过自制工具进行捕捉;另外在蝙蝠活动聚集地待夜色较暗时利用抄网捕捉飞行高度较低的个体(岑业文,彭红元,2010;黄继展等,2013)。采集到的标本进行体质量、体长与前臂长的测量与记录,根据Smith和解焱(2009)进行物种鉴定,对难以野外当场确定的样本带回实验室作进一步研究比对。每个种类蝙蝠均保留对应的浸制标本。 2 结果 2.1 蝙蝠种类与数量
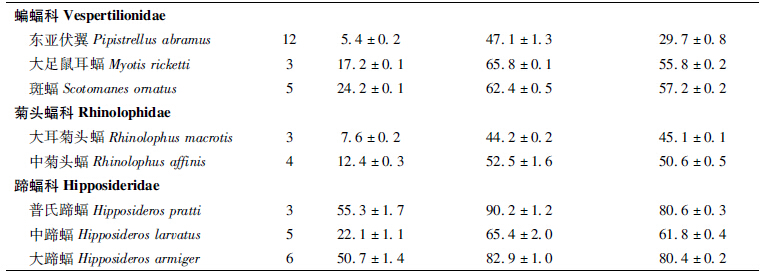
调查共采集到蝙蝠样本8种,分属3科5属,其中蝙蝠科3属3种,菊头蝠科1属2种,蹄蝠科1属3种。8种蝙蝠的分布型中,季风型、古北型、南中国型各1种,其余5种均为东洋型(62.5%)。种群规模上占优势的为东亚伏翼Pipistrellus abramus与大蹄蝠H. armiger,前者数量超过600只,后者200~300只;其次为中菊头蝠R. affinis与中蹄蝠H. larvatus,数量在100~200只之间;数量较少的为普氏蹄蝠H. pratti、大耳菊头蝠R. macrotis,数量在50~100只之间;大足鼠耳蝠M. ricketti与斑蝠Scotomanes ornatus最为稀少,前者数量不超过50只,后者则更是少于20只。调查发现采集的蝙蝠种类、分布型及种群规模见表 1,各种蝙蝠样本采集数量及主要测量数据见表 2。
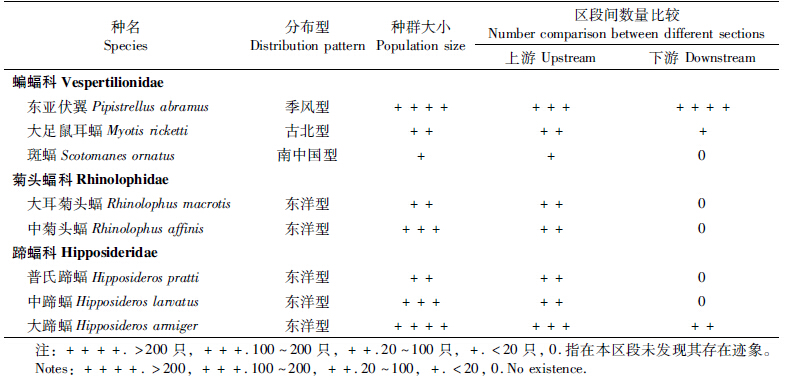
|
将潇水零陵段进入城郊的末段10 km划为下游区,其余为上游区。两段的生态景观及人类活动强度等存在明显差异,主要是由于下游进入城郊,相对上游成片植被覆盖减少,建筑物增加,人类活动更为活跃。调查发现上、下游蝙蝠的种类与种群规模存在明显区别。表 1中等级差异反映出不同区段的数量差异,数字0表示此区段未发现某种蝙蝠存在迹象。从表 1可以看出,8种蝙蝠中,上、下游段均有发现的种类只有3种,分别是东亚伏翼、大足鼠耳蝠与大蹄蝠,其余均只在上游段发现;上、下游段均存在的3种蝙蝠中,东亚伏翼的种群数量下游段大于上游段,其余2种则是上游段较多。 2.3 蝙蝠栖息地类型
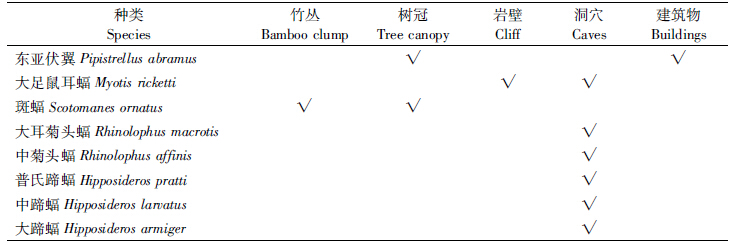
潇水零陵段沿岸的蝙蝠栖息场所(不活动时的休息和居住环境)集中在竹林、树冠、岩壁、洞穴、建筑物等几种类型。竹林主要是枝叶浓密的丛生慈竹丛;树冠主要是叶片密集隐蔽性较好的桂花、广玉兰等树种或一些高大乔木的树冠;岩壁主要是沿河陡峭石壁的狭缝;洞穴包括自然形成或人工作业遗留的洞穴,但调查未发现大规模蝙蝠群体共存的洞穴;建筑物则主要是位置僻静、年代较久的老建筑(如柳子庙等古建筑和老旧民居等)的屋檐、墙缝及瓦片间。除洞穴与建筑物外,其他栖息场所在冬季未见蝙蝠迹象。各蝙蝠种类的栖息类型见表 3,其中东亚伏翼在夏季有停留到树冠歇息现象(在朝阳公园及湖南科技学院校园等处共观察记录到14次,其在树冠停留过程中会偶尔发出人耳可听见的低频声)。
观察发现,区域内蝙蝠主要集中在几处固定地点活动。活动聚集点的上空蝙蝠数量多、密度大。这些活动聚集点一般处于临近河道或人工湖(如怀素公园与朝阳公园内人工湖)等有较大水域面积且周边密集覆盖植被的上空。观察中并未发现蝙蝠活动区域明显远离人类活动场所或聚集地,但人类活动的强度和密度会明显影响蝙蝠活动的高度,且破坏性的人类活动如砍伐、拆迁、爆破等会对蝙蝠的生存造成严重干扰,蝙蝠会转移他处躲避危险。 2.5 不同栖息地类型蝙蝠年活动周期不同
不同种类蝙蝠一年中外出活动的开始与结束日期不同。在洞穴冬眠的蝙蝠较藏于建筑物内过冬的种类外出活动时间晚。如东亚伏翼在3月中下旬偶有单只或少数几只在高温日(>18 ℃)不成规律的外出活动迹象,且活动范围不远离栖息地,而这期间洞穴型蝙蝠未见出洞活动。当冬季即将来临时,洞穴型蝙蝠外出活动的结束日期也较房屋型蝙蝠早。11月上旬所有洞穴型蝙蝠不再外出活动时,东亚伏翼还有零星活动迹象。因此,总体上洞穴型冬眠蝙蝠较房屋型冬眠蝙蝠年活动周期短。 3 讨论
世界各地除少数极端气候或环境地区外均有蝙蝠活动的迹象。在我国,蝙蝠同样呈种类多、分布广等特征(张荣祖等,1997)。加上近年发现的新纪录,我国现存蝙蝠种类已超过130种,但不同地区由于气候与环境的差异,蝙蝠种类及种群规模不尽相同,在国内不同地区开展的蝙蝠资源调查结果各不相同很好地证明了这一点(黎道洪,罗泰昌,2002;岑业文,彭红元,2010;周江,杨天友,2010;黄继展等,2013)。潇水是湘江的重要支流,其零陵段沿岸林木成荫、四季常青。本次调查记录到分属多个科属的8种蝙蝠,且种群整体规模可观,说明潇水零陵段沿岸适合蝙蝠的生存。调查发现的所有蝙蝠种类均在湖南省发现和报道过,未有湖南省新纪录发现,但斑蝠在永州的记录是首次被报道。需注意的是,斑蝠与大足鼠耳蝠数量稀少,应该引起重视,对其进行持续关注与跟踪保护。
蝙蝠与生境存在密切而复杂的关系。一方面蝙蝠依赖所处生境,生物因素(如种间竞争、食物丰度、天敌威胁强度等)与非生物因素(如区域内温度、湿度、光照、噪声等)均会影响蝙蝠对环境的选择(Alcock,1998)。另一方面,蝙蝠也能一定程度上适应所处生境并在行为学上体现出来,如蝙蝠会调节静止频率来适应不同的气候条件和湿度(江廷磊,2010)。潇水河零陵段沿岸集水体(河流与人工湖泊)、植被(灌木、乔木、杂草、慈竹丛、作物等)、农田、菜地、民居及古建筑等多种景观,这给栖息在区域内的蝙蝠提供了良好的栖息与繁殖场所、丰富的食物和便捷的水源。但不同种类的蝙蝠偏好不同类型的生境,如大蹄蝠与菊头蝠多喜灌木林生境,而蝙蝠科种类多喜农田、旱地生境,人们认为这与不同类型生境中蝙蝠的捕食对象——昆虫的种类构成差异有密切关系(周江,杨天友,2010)。潇水河零陵段下游进入城郊,民居与各类其他建筑及农地等增加,而植被覆盖率减少,且人类活动强度增加,这与上游存在明显差异。因此上、下游两段间蝙蝠种类与数量存在差异(表 1)。观察发现,水体与植被衔接处及附近上空的昆虫数量与密度更高,而栖息在沿岸的蝙蝠除大足鼠耳蝠食鱼外均以昆虫为食,因此蝙蝠在水体与植被交界处更易获得食物与水源,从而在这一区域形成活动聚集地。
虽然栖息于某一区域的蝙蝠往往形成较规律的活动节律,包括年活动周期、日活动时间区间、群体规模随月份动态变化等(Meyer et al,2004;廖阳等,2014)。但我们发现,不同蝙蝠种类在一年中外出活动的起始与结束日期存在差异,洞穴内过冬的蝙蝠较在建筑中过冬的蝙蝠年活动周期短。这可能与2个因素有关,一是不同栖息地类型的蝙蝠对低温的耐受能力不同,房屋型蝙蝠的低温耐受能力较洞穴型的更强;二是所处环境的差异导致个体对外界环境温度与光照的变化敏感度不同,房屋型蝙蝠更易觉察和感知到外界环境及气候因子的变化,而气候因子及其导致的蝙蝠食物种类与丰度的差异已被证实为影响蝙蝠地理分布及活动规律的重要因素(李玉春等,2005;廖阳等,2014)。当然,这需要更进一步的研究结果来证实。
蝙蝠对农林生态平衡具有积极效应,因此对蝙蝠资源进行保护具有十分重要的意义,这要求我们首先要对其生存环境进行保护。然而国内蝙蝠生存环境被人类破坏(如过度的旅游资源开发、森林砍伐等),导致蝙蝠种类与数量急剧下降的事件时有发生(石红艳等,2006;Zhang et al,2009)。从本次调查结果可知,潇水永州零陵段沿岸的蝙蝠物种多样性较为丰富,与区域内良好的生态环境密不可分。在永州近年建设旅游型城市背景下,进行潇水沿岸景区开发与城市建设时应增强民众的环境保护意识,为蝙蝠等野生动物留下良好的栖息环境尤为重要。
| 岑业文, 彭红元. 2010. 广西玉林市翼手类多样性初步调查[J]. 四川动物, 29(5): 609-612. |
| 韩宝银, 贺红早. 2012. 大蹄蝠的食物组成及对森林虫害发生的影响[J]. 安徽农业科学, 40(26): 12884-12885. |
| 黄继展, 谭梁静, 杨剑, 等. 2013. 澳门翼手类物种多样性调查[J]. 兽类学报, 33(2): 123-132. |
| 江廷磊. 2010. 蝙蝠回声定位声波地理变化的影响因素及对噪声干扰的反应[D]. 哈尔滨: 东北师范大学. |
| 江廷磊, 刘颖, 冯江. 2008. 中国翼手类一新纪录种[J]. 动物分类学报, 33(1): 212-216. |
| 黎道洪, 罗泰昌. 2002. 云南石林地区岩溶洞穴动物物种多样性初步研究[J]. 贵州师范大学学报: 自然科学版, 20(1): 1-5. |
| 罗丽, 卢冠军, 罗金红, 等. 2011. 湖南省蝙蝠新纪录——大足鼠耳蝠[J]. 动物学杂志, 46(2): 148-152. |
| 廖阳, 闫荣玲, 李金亮, 等. 2014. 永州城郊蝙蝠晚间活动节律初探[J]. 四川动物, 33(2): 224-228. |
| 李玉春, 蒙以航, 张利存, 等. 2005. 中国翼手目地理分布的环境因子影响分析[J]. 动物学报, 51(3): 413-422. |
| 李艳丽, 张佑祥, 刘志霄. 2012. 湖南省翼手目新纪录——大耳菊头蝠[J]. 四川动物, 31(5): 825-827. |
| Smith AT, 解焱. 2009. 中国兽类野外手册[M]. 长沙: 湖南教育出版社. |
| 石红艳, 刘昊, 吴毅, 等. 2006. 四川绵阳洞栖蝙蝠多样性及受胁现状[J]. 四川动物, 25(1): 128-130. |
| 谭敏, 朱光剑, 洪体玉, 等. 2009. 中国翼手类新记录——小蹄蝠[J]. 动物学研究, 30(2): 204-208. |
| 王应祥. 2003. 中国哺乳动物种和亚种分类名录与分布大全[M]. 北京: 中国林业出版社: 27-60. |
| 吴毅, 杨奇森, 夏霖, 等. 2004. 中国蝙蝠新记录——马氏菊头蝠[J]. 动物学杂志, 39(5): 109-110. |
| 吴毅, 易祖盛, 江海声, 等. 2002. 广东英德石门台自然保护区啮齿类及翼手类物种多样性研究[J]. 广州大学学报: 自然科学版, 1(2): 21-26. |
| 赵辉华, 张树义, 周江, 等. 2002. 中国翼手类新记录——高鞍菊头蝠[J]. 兽类学报, 22(1): 74-76. |
| 张礼标, 巩艳艳, 朱光剑, 等. 2010. 中国翼手目新纪录——马来假吸血蝠[J]. 动物学研究, 31(3): 328-332. |
| 周江, 杨天友. 2010. 贵州省松桃县东部地区翼手目物种多样性[J]. 动物学杂志, 45(2): 52-59. |
| 张劲硕, 张礼标, 赵辉华, 等. 2005. 中国翼手类新纪录——小褐菊头蝠[J]. 动物学杂志, 40(2): 96-98. |
| 张礼标, 张劲硕, 梁冰, 等. 2004. 中国翼手类新记录——小巨足蝠[J]. 动物学研究, 25(6): 556-559. |
| 张荣祖, 金善科, 全国强, 等. 1997. 中国哺乳动物分布[M]. 北京: 中国林业出版社: 23-55. |
| Alcock J. 1998. Animal behaviour: an evolutionary approach[M]. Sunderland, Massachusetts, Sinauer Associates Inc Publishers: 555-599. |
| Altringham JD. 1996. Bats-biology and behaviour[M]. Oxford: Oxford University Press: 128-236. |
| Meyer CFJ, Schwarz CJ, Fahr J. 2004. Activity patterns and habitat preferences of insectivorous bats in a west African forest-savanna mosaic[J]. Journal of Tropical Ecology, 20(4): 397-407. |
| Wilson DE, Reeder DM. 2005. Mammal species of the world: a taxonomic and geographic reference (3rd ed)[M]. Baltimore: Johns Hopkins University Press: 312-529. |
| Wu Y, Motokawa M, Harada M. 2008. A new species of the horseshoe bat of the genus Rhinolophus from China (Chiroptera: Rhinolophidae)[J]. Zoological Science, 25(4): 438-443. |
| Wu Y, Thong VD. 2011. A new species of Rhinolophus (Chiroptera: Rhinolophidae) from China[J]. Zoological Science, 28(3): 235-241. |
| Zhang LB, Zhu GJ, Jones G, et al. 2009. Conservation of bats in China: problems and recommendations[J]. Oryx, 43(2): 179-182. |
| Zhang JS, Han NJ, Jons G, et al. 2007. A new species of Barbastella (Chiroptera: Vespertilionidae) from north China[J]. Journal of mammalogy, 88(6): 1393-1403. |
| Zhou ZM, Guillén-Servent A, Lim BK, et al. 2009. A new species from southwestern China in the Afro-Palearctic lineage of the horseshoe bats (Rhinolophus)[J]. Journal of Mammalogy, 90(1): 57-73. |