 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 赵俊松, 李廷野, 房以好, 肖文, 张淑霞
- ZHAO Junsong, LI Tingye, FANG Yihao, XIAO Wen, ZHANG Shuxia
- 云南大理古城3种繁殖鹭类的巢空间隔离
- Nest Space Partitioning in One Three Mixed-species Heronry in Dali Old City, Yunnan
- 四川动物, 2015, 34: 357-363
- Sichuan Journal of Zoology, 2015, 34: 357-363
- 10.3969/j.issn.1000-7083.2015.03.006
-
文章历史
- 收稿日期:2014-07-28
- 接受日期:2014-12-05






2. 西南林业大学, 昆明650200;
3. 大理学院东喜玛拉雅资源与环境研究所, 云南大理671003
2. College of Southwest Forestry University, Kunming 650200, China;
3. Institute of Eastern-Himalaya Biodiversity Research, Dali University, Dali, Yunnan Province 671003, China
动物集群繁殖可能是由于种群中多数个体对少数个体主动接近所形成的(Danchin et al,1998)。鸟类集群繁殖可以使其受益于被捕食风险、食源地等信息的交换和共享(Ward & Zahavi,1973;Roberts,1996)。集群繁殖的鹭类通过筑巢活动在时间或空间上的隔离来减少物种间的竞争压力。例如,多个鹭类集群繁殖地的研究发现混群繁殖的鹭类在进入繁殖活动时间上普遍存在非同步性(Burger,1978a;Gopi & Pandav,2011)。
鹭类在巢址选择上的空间分化分为水平方向和垂直方向。在水平方向上,多数基于生态位宽度理论(戈峰,2010)的研究认为混群繁殖鹭类通常具有明显的水平分布特征,这与各物种的觅食方向、迁入顺序和树种选择有关(伍烈等,2001;叶芬等,2006;王维奎等,2008;谭飞等,2013)。关于巢位垂直方向的空间隔离研究较多,Burger(1978a,1979)认为在同质林(乔木物种组成单一的树林)中,体型大小或体质量与巢位高度呈正相关。Burger的这一观点得到多个集群繁殖地(包括异质林)研究的支持(Fasola & Alieri,1992;Naugle et al,1996;文祯中等,1998;Parejo et al,1999;叶芬等,2006;Ashoori & Barati,2013),虽然鹭类对异质林中不同树种的选择可以影响鹭类巢的垂直分层(Scherer et al,2014)。朱曦等(1998)的研究发现混群繁殖物种中巢位最高的物种并不是个体最大的夜鹭Nycticorax nycticorax,而是种群数量更多的白鹭Egretta garzetta。上述研究说明具体繁殖地的植物多样性、混群物种构成都可能是影响集群繁殖鹭类巢位垂直分层的重要因素;也说明了研究结论具有地点特异性,根据某研究地得出的结论不能外推至其他研究地。上述有关集群鹭巢空间隔离的研究多基于自然繁殖地开展,对位于城镇区域人为干扰较大的繁殖地开展的类似研究尚未见报道。
在自然繁殖地,人为干扰活动少、甚至没有,繁殖地周围觅食地的质量和数量是决定繁殖地的首要因素;与自然繁殖地不同,在人为干扰严重地区繁殖的鹭类面临频繁的人为干扰,筑巢周围环境条件较为恶劣,安全的繁殖地是优先选择,觅食地并非限制因子(Kelly et al,1993;Custer et al,2004)。中国长江流域(Fasola et al,2004)、珠江流域(Wong et al,2004)和海南省(Liang et al,2006)部分鹭类的集群繁殖地位于居民区或人为干扰严重的公园、村庄附近的风水林中,并面临人类威胁,例如人为捕捉幼鸟或采集鸟卵,而欧美国家的鹭类繁殖地多远离人为干扰(Fasola et al,2004),因此中国的鹭类及其繁殖地急需保护。并且在城市集群繁殖的鹭类会因大量鸟粪带来异味、传播潜在疾病、破坏古木而造成人鸟活动冲突(杨月伟,2000;王文林等,2004),在城镇区域繁殖地开展鹭类繁殖研究,将有助于减缓鹭类与人类活动的冲突,保护繁殖鹭类,同时维持良好的城市环境。
对城镇区域鹭类繁殖地的研究报道见于山东省曲阜市(杨月伟,2000)和河南省郑州市(王文林等,2004),均对市区繁殖的鹭鸟生态习性进行了观察,但是对市区的繁殖鹭类巢空间隔离研究尚未见报道。与自然环境繁殖地相比,在城镇区域繁殖地中,繁殖鹭类面临更大的巢址选择压力,为探索城镇区域繁殖地中集群繁殖鹭类的巢空间分布机制,本研究于2012年在云南大理古城市区同质林繁殖地对夜鹭、白鹭、牛背鹭Bubulcus ibis繁殖群体的鹭巢进行了直接计数与定位,研究了混群鹭巢在水平、垂直方向和巢距主干距离上的空间分布特征,旨在揭示仅由这3种鹭类组成的繁殖群体在城镇区域繁殖地中的种间竞争与共存机制。 1 研究地区与研究方法 1.1 研究地概况
研究区域位于云南省大理市古城(大理镇)解放军第六十医院北门口处的圆柏林中(25.70°~25.70°N,100.16°~100.59°E)(图 1),北邻乡镇公路,南邻居民楼。圆柏林为狭长形林带,东西长123 m,南北长17 m,面积2091 m2(图 1),林带被人工修筑的围墙和古城墙所包围,无人进入,形成封闭空间。

|
| 图 1 2012年云南大理古城3种混群繁殖鹭类繁殖地区位与示意图 Fig. 1 Breeding site of the three mixed-species heronry in Dali old city,Yunnan province in 2012 |
研究区域属于北亚热带气候,气候温和,年均气温14~16 ℃,全年多大风,多为西南风(大理州苍山保护管理局,2008)。区域内的树木主要为圆柏Juniperus chinensis,圆柏树木高大整齐,树高较均一,不超过20 m,枝杈繁多,郁闭度较高,加上林下相对封闭的空间,可以为鹭类提供闹市中人为干扰较少的安全环境。
研究区域距洱海的湖滨带3~4 km,同时周围还分布有稻田、鱼塘、小溪等,这些觅食场所为繁殖鹭类提供了丰富的食物来源。据当地居民回忆,该繁殖地至今已经被连续利用7年。2014年春季,医院对该片圆柏林进行修枝,缩小了树木冠幅,迫使鹭类放弃了该片繁殖地。 1.2 研究方法 1.2.1 巢位坐标确定
研究区域为狭长形(图 1),东西向长,南北向窄。正式调查之前,以1 m为单位用皮尺在研究区域边缘加以标记,使研究区域内的树对应相应坐标。研究区域的西北角标记为原点(0,0),以东西方向为X轴,南北方向为Y轴;向东最远记为(120,0),向南最远记为(0,17)。正式调查时,通过预先做好的标记确定鹭巢所在的水平坐[KG(2x]标;以坐标(X,Y)表示水平方向上分布。东西水平方向上以20 m为单位,将0~120 m划分为6个资源等级;南北水平方向上以3 m为单位,将0~17 m划分为6个资源等级。 1.2.2 调查方法
2012年2月初开始每隔1~2 d下午16∶00—19∶00到繁殖地观察有无鹭类在此栖息的痕迹(包括林中的鹭类及林下粪便),判断鹭类迁入该繁殖地的时间,共监测10次;在确定多数鹭类个体完成筑巢后,在繁殖地以北外围公路和繁殖地以南居民楼小道沿路调查记录每个巢的水平坐标、垂直高度和巢距主干的距离。垂直高度以5 m为单位,将0~20 m划分为4个资源等级;巢距主干的距离以0.5 m为单位,将0~2.5 m划分为6个资源等级(含0 m)。持续监测41次后最终确定3种鹭类分别在该繁殖地的营巢分布与位置;待繁殖地内鹭巢位置与巢主物种确定后,每2天监测一次,直至2012年5月末所有鹭类离巢而去,共监测18次。 1.2.3 数据处理
鹭类种群生态位宽度(B),采用基于Shannon-Wienner多样性指数的生态位宽度(戈峰,2010),公式如下:
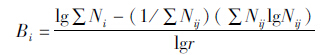
其中,Bi=i物种的生态位宽度;Nij=i物种利用j资源等级的数值;r=生态位资源等级数(在本研究中,东西、南北水平方向上均为6,垂直方向上为4,巢距主干的距离为6);Bi的取值范围是0~1。
种群生态位重叠(Cih),即生态位相似性比例,采用Schoener(1968)的公式计算。
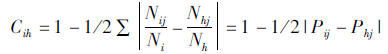
其中,Cih=i物种和h物种之间的比例相似性;Nij=i物种在j资源等级中出现的数值;Ni=i物种在所有资源等级中的数值;Nhj=h物种在j资源等级中出现的数值;Nh=h物种在所有资源等级中的数值;Pij和Phj分别是i物种和h物种在j资源等级中的出现数占各自个体总数的比例。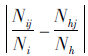 为绝对值,反映2个物种之间利用资源比例之差。0≤Cih≤1,0表示完全不重叠,1表示百分之百的重叠。
为绝对值,反映2个物种之间利用资源比例之差。0≤Cih≤1,0表示完全不重叠,1表示百分之百的重叠。
使用Pearson's Chi-squared test分别检验3种鹭类在东西水平方向、南北水平方向、垂直方向和巢距主干距离上不同资源等级间比例分布差异是否有统计学意义,检验在统计软件R 2.15.0中完成。 2 结果与分析 2.1 群落组成
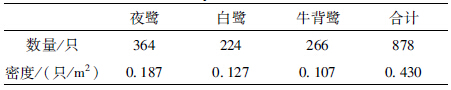
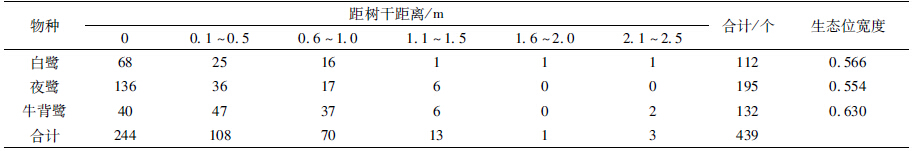
据统计,2012年在大理古城圆柏林内繁殖栖息的鹭类有夜鹭、白鹭和牛背鹭3种,巢数分别为195个、112个和132个。每种鹭类的个体总数按每巢1对鸟计算(朱红星等,1994)。2012年在大理古城圆柏林内繁殖栖息的鹭类总数量为878只,密度0.430只/m2,夜鹭为优势种(表 1)。
|
3种鹭类在迁入与迁出时间上表现出非同步性(图 2),夜鹭迁入、迁出均最早;白鹭紧跟夜鹭迁入和迁出;而牛背鹭迁入最晚,迁出也最晚,滞后夜鹭和白鹭1个月左右。3种鹭类在时空上的迁入情况也是完全不同的。2月末,极少量的夜鹭和白鹭首先迁入,但并未占据研究区域东部的核心区域,而是选用营巢地的旧巢加以修补利用,同时还伴有白鹭和夜鹭之间为了旧巢相互驱逐追打的现象;3月上、中旬,夜鹭开始分批次陆续迁入,并占据繁殖地东部的核心区域,然后向西扩展,同时伴有抢夺白鹭已经占用的旧巢的现象;3月中、下旬,白鹭开始分批次陆续迁入,巢域占据林带中下层;牛背鹭迁入最晚(4月初),其巢域大都分布在繁殖区域的外围,分布区域比白鹭更广。在该繁殖区域内共有白鹭-夜鹭、白鹭-牛背鹭、夜鹭-牛背鹭、白鹭-夜鹭-牛背鹭4种混群形式。还有全是白鹭、全是夜鹭或全是牛背鹭营巢在同一株树上的情况。鹭群在该繁殖地栖息繁殖55~60 d,5月下旬全部离巢。

|
| 图 2 3种鹭类在大理古城繁殖地的停留时间 Fig. 2 Stay duration of three species of egrets in Dali old city breeding site |
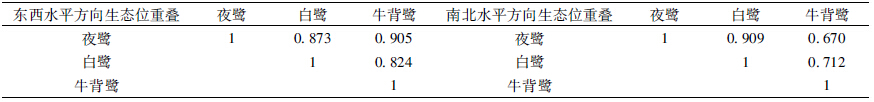
3种鹭类在东西水平方向上的分化不显著(X2=10.2634,df=10,P=0.4177)。生态位宽度上,牛背鹭最大,为0.912;夜鹭最小,为0.750(表 2)。夜鹭和白鹭的巢均在研究区域距西侧围墙100.1~120.0 m之间最多,分别有57个和40个,分别占其巢总数的29.2%和35.7%,而牛背鹭的巢在距西侧围墙80.1~100.0 m处及100.1~120.0 m处均为34个,占其巢总数的25.8%。夜鹭和牛背鹭在东西水平方向上的生态位重叠值最大,为0.905(表 4)。
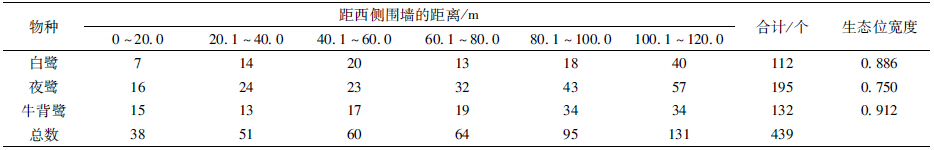
|
3种鹭类在南北水平方向上的分化不显著(X2=4.9234,df=8,P=0.7657)。生态位宽度上,夜鹭最大,为0.858;白鹭最小,为0.809(表 3)。3种鹭类的巢分布最大值均在距北侧公路0~3 m处,其中白鹭为32个,夜鹭56个,牛背鹭47个,分别占其巢总数的28.6%、28.7%和35.6%。夜鹭和白鹭在南北水平方向上的生态位重叠值最大,为0.909(表 4)。
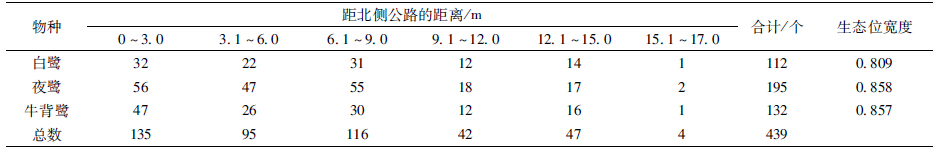
|
|
3种鹭类在垂直方向上的分化显著(X2=114.6679,df=2,P<0.01)。生态位宽度上,白鹭最大,为0.584;夜鹭最小,为0.288(表 5)。夜鹭和牛背鹭的巢均在中高层(巢高10.1~15.0 m)最多,分别有175个和96个,分别占其巢总数的89.7%和72.7%,而白鹭的巢在中低层(巢高5.1~10.0 m)最多,有68个,占其巢总数的60.7%。夜鹭和牛背鹭在垂直方向上的生态位重叠值最大,为0.819(表 6)。
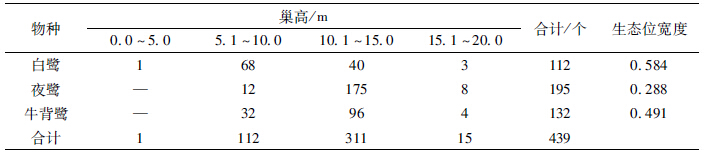
|
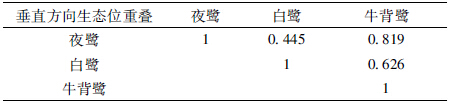
|
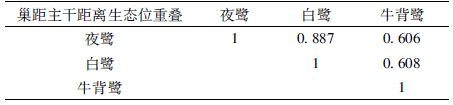
3种鹭类在巢距主干距离上的分化显著(X2=53.5004,df=6,P<0.01)。生态位宽度上,牛背鹭最大,为0.630;夜鹭最小,为0.554(表 7)。夜鹭和白鹭的巢多数分布在主干上(距树干距离为0 m),分别有136个和68个,分别占其巢总数的69.7%和60.7%;而牛背鹭的巢多数在侧枝上,在主干上的巢最少,仅有40个,占其巢总数的30.3%。夜鹭和白鹭在巢距主干距离的生态位重叠值最大,为0.887(表 8)。
|
|
本研究中3种鹭巢在东西和南北水平方向上均无显著分化,该结果与国内多数基于自然繁殖地的研究结果不同(伍烈等,2001;叶芬等,2006;王维奎等,2008;谭飞等,2013)。伍烈等(2001)研究的厦门白鹭自然保护区的繁殖地面积为1.8 km2;叶芬等(2006)研究的广西防城万鹤山鹭鸟自然保护区繁殖地面积为1 hm2;王维奎等(2008)研究的四川南充太和白鹭自然保护区繁殖地面积为1.23 hm2;谭飞等(2013)研究的福建漳江口红树林湿地自然保护区繁殖地面积为103.75 hm2;本研究中位于大理古城市区的繁殖地面积仅为0.209 hm2,较小的繁殖地面积可能是造成3种鹭巢在水平方向不易产生分化的原因。
本研究中3种鹭巢的垂直分化显著,夜鹭占据高位巢的比例最大。与白鹭和牛背鹭相比,夜鹭的巢分布较为集中,89.7%的巢分布在中高位林层。Burger(1978a,1979)认为在同质林中体型大小或体质量与巢位高度呈正相关。夜鹭为3种鹭类中体质量最大的(文祯中等,1998; Parejo et al,1999),因此其在种间竞争时占有绝对优势,在我们的实际观察中也发现夜鹭对异种入侵者有明显的驱赶打斗,且占上风。高位巢的繁殖成功率高可能是夜鹭占据高位巢的原因,因为高位巢具有较多优点,例如更容易发现捕食者、容易进入巢中、避免与来自地面的捕食者相遇(Burger,1979)。Park等(2011)发现6种鹭类的繁殖成功率与巢高呈正相关,夜鹭的实际巢高高于体质量-巢高正相关理论预期的巢高;Ashoor和Barati(2013)的研究也表明对于夜鹭的不同个体,占据高位巢的个体繁殖成功率也较高。另外,迁入顺序对鹭类巢的垂直分布也有影响(Burger & Gochfeld,1990),最早迁入者往往占据最有利的巢位,本研究中夜鹭是3种鹭类中最早迁入的,因此综合体质量和迁入顺序两方面的因素,89.7%的夜鹭巢分布于林层的中高处(10.1~15 m)。
本研究中夜鹭巢集中分布于筑巢树木主干上,与自然繁殖地中夜鹭巢远离筑巢树木主干的结果不同。叶芬等(2006)对广西防城万鹤山鹭鸟保护区7种混群鹭巢的空间隔离研究表明,与其他6种混群鹭类相比,夜鹭的巢距离主干最远。Kim和Koo(2009)的研究表明鹭巢与树主干的距离对夜鹭的繁殖成功率有较大影响,远离树干的夜鹭巢极易暴露在天敌下,从而使夜鹭的繁殖成功率小于其他共同繁殖的鹭类物种。在城市繁殖地较大的巢址选择压力下,夜鹭的巢靠近主干可能是提高自身繁殖成功率的策略,繁殖地之间的差异是否为夜鹭巢位改变的原因,有待更深入的研究。
在本研究中,没有任何2种鹭类的巢在4个空间方向上发生明显重叠,例如夜鹭和牛背鹭在东西水平和垂直方向有最大的生态位重叠,分别为0.925和0.819,而夜鹭和白鹭在巢距主干距离和南北水平方向,具有最大的生态位重叠,分别为0.887和0.909,说明3个物种在较大的种间竞争压力下,种间巢位的空间生态位分化较为精细。 3.2 云南大理古城市区的繁殖地与鹭类组成特点
树巢型鹭类群落结构多样性受栖息地结构、植物物种多样性、林木的水平和垂直层次复杂性的影响(朱曦,2005)。大理古城的鹭类群落仅由3个物种组成,且总数量不大,这与该繁殖地面积小、植物物种组成单一有直接关系。如果外界条件不改变,鹭类对集群繁殖地有高度的忠诚性,一般会连续利用适宜的繁殖地(Kelly et al,2007)。大理古城的繁殖地仅在近7~8年来开始被利用,说明该繁殖地并不是鹭类原有的繁殖地,可能是由于原有繁殖地的丧失或气候变化而导致的鹭类营巢地迁移,因为鹭类的繁殖活动会受到繁殖季节前降雨、气温变化及人类破坏活动、区域尺度上的种群变化等多因素的影响(Kelly et al,2007;Fasola et al,2010;Kelly & Emiko,2014)。
与其他鹭类相比,牛背鹭、夜鹭和白鹭3种鹭类混群筑巢的形式多样,活动的范围和强度最大,能够适应更广泛的栖息环境(叶芬等,2006)。本繁殖地夜鹭、牛背鹭和白鹭的繁殖混群方式,与中纬度地区西班牙西南部巴达霍斯市(Parejo et al,1999)、伊朗南里海地区(Ashoori & Barati,2013)报道的一致,但是本研究地的牛背鹭为留鸟(杨岚,1995),而非夏候鸟。非留鸟的牛背鹭行为更为激进,从而使其巢位发生变化(Burger,1978b)。
大理古城地处旅游区,我们发现的繁殖地紧邻公路和居民楼,人为干扰极为严重,鹭类偏偏选择这一城区繁殖,反映出近年来该地区的鹭类繁殖压力可能正在增加。2014年该繁殖地的树木大幅修枝又使这些繁殖鹭类放弃了对该繁殖地的利用。由于水稻田的分布与营树巢的鹭类繁殖地密切相关(Toureng et al,2004),因此,保留大理洱海流域自然水体附近的水稻田等湿地及高大乔木林,禁止对已知鹭类繁殖地树木大幅修枝,将有利于保护该地区的繁殖鹭类种群。
| 大理白族自治州苍山保护管理局. 2008. 苍山志[M]. 昆明: 云南民族出版社: 35-43. |
| 戈峰. 2010. 现代生态学(第二版)[M]. 北京: 科学出版社: 318-332. |
| 谭飞, 林英华, 张明海. 2013. 福建漳江口繁殖期鹭科鸟类觅食及巢位空间生态位分离[J]. 野生动物, 34(2): 79-83. |
| 王维奎, 周材权, 龙帅, 等. 2008. 四川南充太和鹭类鸟类群落空间生态位和种间关系[J]. 四川动物, 27(2): 178-182. |
| 王文林, 张丽琴, 李长看, 等. 2004. 郑州鹭类的生态习性[J]. 河南科学, 22(6): 792-795. |
| 文祯中, 王庆林, 孙儒泳. 1998. 鹭科鸟类种间关系的研究[J]. 生态学杂志, 17(1): 27-34. |
| 伍烈, 陈小麟, 胡慧娟, 等. 2001. 厦门白鹭自然保护区鹭类繁殖的空间分布[J]. 厦门大学学报: 自然科学版, 40(4): 979-983. |
| 杨岚. 1995. 云南鸟类志(上卷·非雀形目)[M]. 昆明: 云南科技出版社: 102. |
| 杨月伟. 2000. 山东曲阜鹭的生态学研究[J]. 曲阜师范大学学报, 26(3): 80-83. |
| 叶芬, 黄乘明, 李汉华. 2006. 广西防城7种鹭类混群繁殖的空间生态位研究[J]. 四川动物, 25(3): 577-583. |
| 朱红星, 聂继山, 赵耀宗. 1994. 六种鹭鸟混群营巢地的调查[J]. 四川动物, 13(3): 123-124. |
| 朱曦, 章立新, 梁峻, 等. 1998. 鹭科鸟类群落的空间生态位和种间关系[J]. 动物学研究, 19(1): 45-51. |
| 朱曦. 2005. 中国鹭科鸟类研究进展[J]. 林业科学, 41(1): 174-180. |
| Ashoori A, Barati A. 2013. Breeding success of black-crowned night heron (Nycticorax nycticorax), little egret (Egretta garzetta) and cattle egret (Bubulcus ibis) (Aves: Ardeidae) in relation to nest height in the South Caspian Sea[J]. Italian Journal of Zoology, 80(1): 149-154. |
| Burger J. 1978a. The pattern and mechanism of nesting in mixed species heronries[M].// A Sprunt IV, JC Odgen, S Wincker (Eds.): Wading Birds: 45-58. Research Report of National Audubon Society No.7. |
| Burger J. 1978b. Competition between cattle egrets and native north American herons, egrets and ibises[J]. Condor, 80: 15-23. |
| Burger J. 1979. Resource partitioning: nest site selection in mixed-species colonies of herons, egrets and ibises[J]. American Midland Naturalist, 101: 191-210. |
| Burger J, Gochfeld M. 1990. Vertical nest stratification in a heronry in Madagascar[J]. Colonial Waterbirds, 13: 143-146. |
| Custer CM, Suárez SA, Olsen DA. 2004. Feeding habitat characteristics of the great blue heron and great egret nesting along the upper Mississippi River, 1995-1998[J]. Waterbirds, 27(4): 454-468. |
| Danchin E, Boulinier T, Massot M. 1998. Conspecific reproductive and breeding habitat selection: implications for the study of coloniality[J]. Ecology, 79: 2415-2428. |
| Fasola M, Alieri R. 1992. Nest site characteristics in relation to body size in herons in Italy[J]. Colonial Waterbirds, 15: 185-192. |
| Fasola M, Galeotti P, Dai N, et al. 2004. Large numbers of breeding egrets and herons in China[J]. Waterbirds, 27(1): 126-128. |
| Fasola M, Rubolini D, Merli E, et al. 2010. Long-term trends of heron and egret populations in Italy, and the effects of climate, human-induced mortality, and habitat on population dynamics[J]. Population Ecology, 52(1): 59-72. |
| Gopi GV, Pandav B. 2011. Nest space partitioning among colonial nesting waterbirds at Bhitarkanika Mangroves, India[J]. World Journal of Zoology, 6(1): 61-72. |
| Kelly JP, Emiko CT. 2014. Rainfall effects on heron and egret nest abundance in the San Francisco Bay area[J]. Wetlands, 34(5): 893-903. |
| Kelly JP, Etienne K, Strong C, et al. 2007. Status, trend, and implications for the conservation of heron and egret nesting colonies in the San Francisco Bay area[J]. Waterbirds, 30(4): 455-478. |
| Kelly JP, Pratt HM, Greene PL. 1993. The distribution, reproductive success, and habitat characteristics of heron and egret breeding colonies in the San Francisco Bay area[J]. Colonial Waterbirds, 16: 18-27. |
| Kim J, Koo TH. 2009. Nest site selection and reproductive success of herons and egrets in Pyeongtaek Heronry, Korea[J]. Waterbirds, 32(1): 116-122. |
| Liang W, Wong LC, Wong JYP. 2006. Arderid nesting colony survey in Hainan, China[J]. Waterbirds, 29(1): 69-75. |
| Naugle DE, Johnson RR, Meeks WA, et al. 1996. Colonazation and growth of a mixed-species heronry in south Dakota[J]. Colonial Waterbirds, 19(2): 199-206. |
| Parejo D, Sánchez JM, Avilés JM. 1999. Factors affecting the nest height of three heron species in heronries in the south-west of Spain[J]. Ardeola, 46(2): 227-230. |
| Park SR, Kim KY, Chung H, et al. 2011. Vertical nest stratification and breeding success in a six mixed-species heronry in Taeseong, Chungbuk, Korea[J]. Animal Cells and Systems, 15(1): 85-90. |
| Roberts G. 1996. Why individual vigilance declines as group size increase[J]. Animal Behavior, 51: 1077-1086. |
| Scherer JM, Scherer AL, Petry MV. 2014. Vertical nest stratification of four heron species in southern Brazil[J]. Studies on Neotropical Fauna and Environment, 49(1): 66-74. |
| Schoener TW. 1968. The Anolis lizards of Bimini: resource partitioning in a complex fauna[J]. Ecology, 49: 704-726. |
| Toureng C, Benhamou S, Sadoul N, et al. 2004. Spatial relationships between tree-nesting heron colonies and rice fields in the Camargue, France[J]. The Auk, 121(1): 192-202. |
| Ward P, Zahavi A. 1973. The importance of certain assemblages of birds as "Information-centers" for food-finding[J]. Ibis, 115(4): 517-534. |
| Wong LC, Gao Y, Chang H, et al. 2004. A questionnaire survey of ardeid nesting colony distribution in Guangdong, Guangxi and Hainan, south China[J]. Waterbirds, 27(2): 216-223. |