 2015, Vol. 34
2015, Vol. 34扩展功能
文章信息
- 夏万才, 黎大勇, 李艳红, 和鑫明, 钟泰, 项雷, 胡杰
- XIA Wancai, LI Dayong, LI Yanhong, HE Xinming, ZHONG Tai, XIANG Lei, HU Jie
- 云南白马雪山国家级自然保护区响古箐地区鸟类物种多样性垂直分布格局
- Avian Diversity along the Elevational Gradient of Xiangguqing in Baimaxueshan National Nature Reserve, Yunnan Province
- 四川动物, 2015, 34(4): 620-625
- Sichuan Journal of Zoology, 2015, 34(4): 620-625
- 10.11984/j.issn.1000-7083.2015.04.022
-
文章历史
- 收稿日期:2014-10-22
- 接受日期:2015-02-25






2. 西华师范大学珍稀动植物研究所, 四川南充 637009;
3. 白马雪山自然保护区管理局, 云南迪庆 674400
2. Institute of Rare Animals and Plants, China West Normal University, Nanchong, Sichuan Province 637009, China;
3. Baimaxueshan National Nature Reserve, Diqing, Yunnan Province 674400, China
野生动植物种群和生物多样性沿海拔梯度的垂直分布模式和机制是生物多样性和生态学研究中的重要问题,也是生物多样性保护和自然资源管理实践中亟需了解的信息基础(吴永杰,雷富民,2013)。生物多样性垂直分布模式的研究一直是生态学研究的热点(Fu et al.,2006;Brehm et al.,2007;马俊等,2010;吴永杰等,2012;吴永杰,雷富民,2013;李晟等,2014)。目前开展了蚤类(龚正达等,2005)、小型哺乳动物(马俊等,2010;吴永杰等,2012)、有蹄类(李晟等,2014)、鸟类(王庆忠,王大科,1995;Bhatt & Joshi,2011;何中声等,2012;粟通萍等,2012;王斌等,2013; Wu et al.,2013)等类群物种多样性垂直分布的研究。研究鸟类多样性不仅是资源保护的需要,由于鸟类对环境变化的响应是最敏感迅速的,因此,对鸟类多样性的比较能对环境的变动及质量做出客观评价(Mǒrtberg,2001;乌日罕等,2014)。
白马雪山国家级自然保护区位于全球生物多样性热点区域之一的中国西南山地,地处横断山脉中部偏南,是“三江并流”世界遗产地范围内的5个保护区之一,主要保护对象为滇金丝猴Rhinopithecus bieti、红豆杉Taxus chinensis等珍稀濒危野生动植物及其生境。至今有关该保护区鸟类的调查仍然薄弱(云南省林业厅,2003;阿青等,2006;夏万才等,2014)。唐蟾珠(1996)对横断山区的鸟类物种丰富度的垂直分布进行了初步研究。在此基础上,Wu等(2013)对物种丰富度分布格局和影响机制的研究发现横断山地区鸟类的物种丰富度与气候和能量因子关系最为密切。迄今为止,尚未见有对位于保护区南端——响古箐地区鸟类的研究报道。为此,我们于2013年7月—2014年2月对云南白马雪山国家级自然保护区响古箐地区鸟类的多样性和空间分布格局进行了系统调查和研究,以期进一步丰富该区域鸟类多样性的基础资料,并为本地区的生物多样性保护提供重要参考。夏万才等:白马雪山国家级自然保护区响古箐地区鸟类物种多样性垂直分布格局 1 研究地概况
响古箐位于云南省迪庆州维西县塔城镇(27°36′N,99°15′E)境内,白马雪山国家级自然保护区的南端。该研究地东西宽约9 km,南北长约10 km,总面积约90 km2(黎大勇,2010)。响古箐是典型的山地切割地貌,境内最高海拔4100 m左右,最低海拔2400 m左右,相对海拔高差约为1700 m。响古箐所处地属于高原山地季风气候,一年中明显分为旱、湿两季,5月至10月是湿季,降雨集中,湿度大,气温高;11月至翌年4月是旱季,降雨稀少,
空气干燥,气温低。研究地年降雨量1370.7 mm,年平均气温为9.8 ℃(黎大勇,2010)。参考《云南植被》(吴征镒,朱彦丞,1987),将研究区域划分为5种植被类型:云南松林、常绿阔叶林、针阔混交林、寒温性针叶林、高山灌丛草甸。 2 研究方法 2.1 调查方法
调查期间,根据据保护区内地形、地貌及植被等自然景观特点,共设置了34条样线,每条样线调查4次,调查时采用无距离样线法,以平均2~3 km·h-1的速度行进,使用Olympus 10×42双筒望远镜记录统计样线两侧和前方鸟类的种类及数量。结合《中国鸟类图鉴》(约翰·马敬能等,2000)、《云南鸟类志》(杨岚等,1994,2004)进行野外识别,并参照《中国鸟类分类与名录分布(第二版)》(郑光美,2011)进行分类;动物区系和分布型划分依据《中国动物地理》(张荣祖,1999)。 2.2 数据处理 2.2.1 多样性指数H′
采用Shannon-Wiener指数计算:
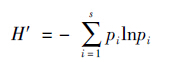
其中,s为物种的种类数,pi为第i种物种的个体数量占全部物种个体数量的比例。
2.2.2 均匀度J用Pielou均匀度指数J表示(Pielou′s evenness index):
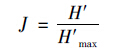
其中H′为香农指数,H′max是H′的最大值:
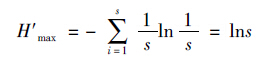
采用优势度指数公式计算(栾晓峰等,2003):
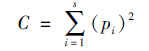
根据Berger-Paker优势度指数(韩联宪等,2014):
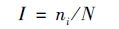
I≥0.05为优势种;0.005≤I<0.05为常见种;I<0.005为偶见种。ni为物种i的个体数量,N为全部物种的总个体数量。
数据用Excel处理。

|
| 图 1 云南白马雪山国家级自然保护区响古箐地区地形图和样线位置图 Fig. 1 Study area and survey line along the Xiangguqing in Baimaxueshan National Nature Reserve |
本次调查响古箐地区共记录到鸟类139种,隶属9目36科,属于国家Ⅰ级重点保护的鸟类有红喉雉鹑Tetraophasis obscurus和黑颈长尾雉Syrmaticus humiae 2种;属于国家Ⅱ级重点保护鸟类有白头鹞Circus aeruginosus、雀鹰Accipiter nisus、普通Buteo buteo、红隼Falco tinnunculus、血雉Ithaginis cruentus、白腹锦鸡Chrysolophus amherstiae、楔尾绿鸠Treron sphenura、灰林鸮Strix aluco和白腹黑啄木鸟Dryocopus javensis 9种。其中,中国特有种包括红喉雉鹑、大噪鹛Garrulax maximus、橙翅噪鹛G. elliotii、黄腹山雀Parus venustulus、滇Sitta yunnanensis等13种(雷富民等,2002)。按数量等级分:响古箐地区以白领凤鹛Yuhina diademata、方尾鹟Culicicapa ceylonensis和黑短脚鹎Hypsipetes madagascariensis为优势种,占记录物种数的2.16%,个体数占总个体数的25.28%;常见种42种,占记录物种数的30.22%,个体数占总个体数的50.39%;稀有种94种,占记录物种数的67.62%,个体数占总个体数的21.33%。该地的鸟类构成以留鸟为主,有97种;其他分别为夏候鸟28种,冬候鸟7种,旅鸟7种。从区系上来看,响古箐地区鸟类以东洋界成分为主,有73种,占52.52%;广布种34种,占24.46%;古北界种32种,占23.02%。 3.2 鸟类的垂直分布 3.2.1 云南松林
海拔2400~3000 m,有少许村落、农田夹杂其间,以云南松Pinus yunnanensis为优势树种,常形成纯林,林下分布大白花杜鹃Rhododendron decorum、地檀香Gaultheria forrestii、水红木Viburnum cylindricum等植物。共记录到鸟类7目29科69种,其中东洋界32种,古北界17种,广布种20种。优势种为长尾山椒鸟Pericrocotus ethologus、黑短脚鹎和麻雀Passer montanus;常见种有山斑鸠Streptopelia orientalis、家燕Hirundo rustica、金腰燕H. daurica、白鹡鸰Motacilla alba、树鹨Anthus hodgsoni、黄腹柳莺Phylloscopus affinis等66种。
3.2.2 常绿阔叶林海拔2500~2900 m,分布在阴坡和沟谷的局部地段。以曼青冈Cyclobalanopsis oxyodon为优势树种,林下分布着桦叶荚蒾Viburnum betulifolium、川滇腊树Ligustrum delavayanum等植物。共记录到鸟类7目30科78种,其中东洋界39种,古北界18种,广布种21种。优势种为黑短脚鹎、白领凤鹛和麻雀;常见种有楔尾绿鸠、铜蓝鹟Eumyias thalassina、黄眉柳莺P. inornatus、煤山雀P. ater、黄腹山雀、绿背山雀P. monticolus和滇等47种。
3.2.3 针阔混交林海拔2600~3300 m,针叶树主要有云南铁杉Tsuga dumosa、华山松P. arm and ii等;阔叶树主要有糙皮桦Betula utilis、康藏花楸Sorbus thibetica、吴茱萸叶五加Acanthopanax evodiaefolius、丽江槭Acer forrestii等植物;共记录到鸟类9目34科123种,其中东洋界65种,古北界28种,广布种30种。优势种为白领凤鹛、橙翅噪鹛和方尾鹟;常见种有黑卷尾Dicrurus macrocercus、灰卷尾D. leucophaeus、松鸦G. gl and arius、黄嘴蓝鹊Urocissa flavirostris、蓝额红尾鸲Phoenicurus frontalis、乌鹟Muscicapa sibirica、北灰鹟M. dauurica、棕腹仙鹟Niltava sundara、黄腹扇尾鹟Rhipidura hypoxantha、白喉噪鹛G. albogularis、纹喉凤鹛Yuhina gularis、金眶鹟莺Seicercus burkii、大山雀P. major、黄颊山雀P. spilonotus、黄腹啄花鸟、蓝喉太阳鸟Aethopyga gouldiae、灰头灰雀Pyrrhula erythaca等34种。
3.2.4 寒温性针叶林海拔3300~3900 m:3600 m以下油麦吊云杉Picea complanata为优势树种,3600 m以上以长苞冷杉Abies georgei为优势树种,林下分布杜鹃属、箭竹属Sinarundinaria等植物;共记录到鸟类4目19科45种,其中东洋界18种,古北界16种,广布种11种。优势种为白眉雀鹛Alcippe vinipectus、白领凤鹛、橙翅噪鹛、乌鹟和松鸦;常见种有星鸦Nucifraga caryocatactes、沼泽山雀P. palustris、普通S. europaea、红翅旋壁雀Tichodroma muraria、高山旋木雀Certhia himalayana、普通朱雀Carpodacus erythrinus、曙红朱雀C. eos、白眉朱雀C. thura、藏黄雀Carduelis thibetanus等40种。
3.2.5 高山灌丛草甸海拔3900~4100 m,灌丛为多色杜鹃和金黄杜鹃为主的寒温性灌丛;草甸植物主要有西南鸢尾Iris bulleyaan、矮蒿草Kodresia humilis、珠芽蓼Polgonum vivuparum和尼泊尔酸模Rumex nepalensis等。共记录到鸟类5目13科24种,其中东洋界11种,古北界10种,广布种3种。优势种为白眉雀鹛、大噪鹛和北灰鹟;常见种为小嘴乌鸦Corvus corone、大嘴乌鸦C. macrorhynchos和棕背黑头鸫Turdus kessleri等21种。 3.3 不同生境鸟类的多样性
各类生境中的鸟类多样性由高到低依次为:针阔混交林、常绿阔叶林、云南松林、寒温性针叶林、高山灌丛草甸。整体来看,响古箐地区鸟类多样性随着海拔的上升而缓慢增加,到针阔混交林带达到最大值,然后快速降低。鸟类优势度从高到低依次为:高山灌丛草甸、寒温性针叶林、云南松林、常绿阔叶林、针阔混交林(表 1)。
| 生境类型Habitat type | 物种数 Number of species | 香浓-维纳多样性指数 Shannon-Wiener index | 优势度 Dominance index | 均匀度 Evenness index |
| 云南松林Yunnan pine forest | 69 | 3.762 | 0.036 | 0.691 |
| 常绿阔叶林Evergreen broad-leaved forest | 78 | 3.777 | 0.043 | 0.670 |
| 针阔混交林Mixed broadleaf and conifer forest | 123 | 3.786 | 0.052 | 0.571 |
| 寒温性针叶林Cold conifer forest | 45 | 3.203 | 0.067 | 0.650 |
| 高山灌丛草甸Alpine bush and meadow | 24 | 2.871 | 0.066 | 0.750 |
针阔混交林及以下地区,东洋界鸟类比例随海拔的上升而增加,古北界和广布种所占比例随海拔上升而下降;寒温性针叶林以上地区,古北界鸟类比例随海拔上升而增加,而东洋界与广布种鸟类的比例则无明显的规律。针阔混交林与寒温性针叶林的过渡地带(3200~3400 m)成为了该区域鸟类地理成分的“转折点”(图 2)。

|
| 图 2 不同海拔段东洋界、古北界及广布种鸟类的差异 Fig. 2 Differences among the Oriental,Palearctic and widespread in different elevations |
横断山区作为我国生物多样性最丰富、最集中的地区之一,素有“植物王国”“动物王国”之美誉(云南省林业厅,2003)。响古箐地区位于横断山区中南部,其鸟类资源十分丰富,具有特有种多、多样性高等特点。由于响古箐地区处于中国动物地理区划东洋界和古北界的交汇地带,南北物种在此渗透交流(张荣祖,1999)。但从动物地理成分来看,该区域鸟类以东洋界为主(东洋界物种占52.52%;广布种占24.46%;古北界物种占23.02%),同时也反映出了本地区鸟类区系具有古北界与东洋界物种相互渗透和过渡的特点。 4.2 影响鸟类垂直分布的主要因素
云南白马雪山国家级自然保护区与横断山区的大多数山体一样,山高坡陡,植被的垂直带谱较明显。响古箐鸟类多样性呈现出随着海拔升高,鸟类多样性指数先逐渐增加,到针阔混交林带最大,然后迅速下降的这一特征,其与郑作新等(1963)、谭耀匡和郑作新(1964)、粟通萍等(2012)对鸟类垂直分布的研究结果基本吻合。符合Rahbek(2005)总结出的物种多样性在海拔梯度上具有单峰分布的特征,即在中海拔地区物种多样性最高的结论。按照吴永杰和雷富民(2013)对物种丰富度垂直分布格局模式的划分,本研究地鸟类物种丰富度的垂直分布表现为偏前峰模式。王斌等(2013)认为,鸟类的群落结构与栖息地的气候、地形、地势及相关的植被类型密切相关,即生态系统的多样性决定了鸟类群落结构的多样性。受海拔和地形的影响,食物和隐蔽场所是影响鸟类垂直分布的重要因素。中低海拔带植物种类丰富,空间异质性高,食物充足,特别是混交林生境复杂,具备鸟类栖息的多种生境,导致依赖一定植物生存的鸟类聚集,即鸟类的多样性也相应升高(姜云垒等,2006;闫慧等,2011)。前期研究表明在大的区域尺度上,横断山地区鸟类的物种丰富度与气候和能量因子关系最为密切,植物生产力对鸟类的多样性也具有重要影响(Wu et al.,2013)。能量假说认为植物生产力高,鸟类物种丰富度就越高。本研究中针阔混交林以下地区气候温暖湿润,植被茂盛,鸟类物种丰富度在中低海拔地区最高支持Wu等(2013)发现横断山地区鸟类物种丰富度与能量、气候和增强植被指数关系密切的结论。同时中域效应(mid-domain effect)假说认为:受几何边界的限制,不同物种分布区在该几何区域中间位置重叠程度大,而在边界附近重叠少,从而形成物种丰富度从边界向中心逐渐增加的格局(Colwell & Lees,2000)。显然,中域效应假说不是导致本地区鸟类物种丰富度垂直变化的最主要因素。响古箐地区鸟类丰富度在中低海拔地区最高,还与一些生态和历史的因素有关,如干扰和物种进化时间等。该地人口密度和耕地面积较小,人为干扰影响较小;而同时Wu等(2014)的研究发现中低海拔地区的鸟类比高海拔地区的具有更长的进化时间,这意味着更多的物种分化。综上所述,多因素的叠加和综合导致该地区鸟类在垂直海拔梯度分布上呈现出偏前峰模式。 4.3 造成不同生境类型鸟类地理成分差异的因素
唐蟾珠(1996)认为横断山区鸟类随海拔增高,东洋界物种比例减小,古北界鸟类和特有种鸟类增加,广布种无显著差异。而本研究与此理论不符。Wu等(2013)对横断山地区鸟类的研究发现:鸟类的丰富度与气候和能量因子关系最为密切,与气温年较差成反比,与植物生产力正相关,而面积和空间几何边界限制对鸟类物种丰富度影响较小。针阔混交林与寒温性针叶林过渡的区域成为该区域鸟类地理成分分布的“拐点”。针阔混交林以上区域,气温年较差大,气候相对严苛,植被单一,生产力较低,因而鸟类食物资源匮乏,隐蔽条件差,导致鸟类物种多样性较低。而这种环境并不适应东洋界物种生存,但更适合古北界的物种生存,从而造成鸟类区系组成在高海拔地区出现组成(比例)的差别。
致谢:本研究得到了白马雪山国家级自然保护区维西管理分局、西华师范大学大学生创新项目(42713059)、西华师范大学博士科研启动项目(11B018;12B020)的支持,同时感谢评审专家对本文提出的宝贵意见。| 阿青, 格玛江初, 陈永春. 2006. 白马雪山自然保护区德钦分局辖区鸟类多样性及保护管理对策[J]. 林业调查规划, 31(5): 85-88. |
| 龚正达, 吴厚永, 段兴德, 等. 2005. 云南横断山区蚤类物种丰富度与区系的垂直分布格局[J]. 生物多样性, 13(4): 279-289. |
| 韩联宪, 邓章文, 岩道. 2014. 大理洱海湖滨带鸟类多样性[J]. 动物学杂志, 49(2): 185-194. |
| 何中声, 刘金福, 洪伟, 等. 2012. 泉州市鸟类多样性海拔梯度格局的研究[J]. 中南林业科技大学学报, 32(6): 188-191. |
| 姜云垒, 高玮, 王海涛, 等. 2006. 长白山北坡鸟类多样性研究[J]. 东北师范大学学报: 自然科学版, 38(2): 105-108. |
| 雷富民, 屈延平, 卢建利, 等. 2002. 关于中国鸟类特有种名录的核定[J]. 动物分类学报, 27(4): 857-864. |
| 黎大勇. 2010. 白马雪山自然保护区响古箐滇金丝猴Rhinopithecus bieti活动时间分配、夜宿行为和食性研究[D]. 西安: 西北大学. |
| 李晟, 张晓峰, 陈鹏, 等. 2014. 秦岭南坡森林有蹄类群落组成与垂直分布特征[J]. 动物学杂志, 49(5): 633-643. |
| 粟通萍, 王绍能, 蒋爱伍. 2012. 广西猫儿山地区鸟类组成及垂直分布格局[J]. 动物学杂志, 47(6): 54-65. |
| 栾晓峰, 刘俊峰, 胡忠军, 等. 2003. 上海郊区冬夏季鸟类群落特征比较[J]. 动物学杂志, 38(3): 69-76. |
| 马俊, 吴永杰, 夏霖, 等. 2010. 螺髻山自然保护区非飞行小型哺乳动物垂直多样性调查[J]. 兽类学报, 30(4): 400-410. |
| 谭耀匡, 郑作新. 1964. 云南玉龙雪山鸟类的垂直分布[J]. 动物学报, 16(2): 295-313. |
| 唐蟾珠. 1996. 横断山区鸟类[M]. 北京: 科学出版社. |
| 王斌, 彭波涌, 李晶晶, 等. 2013. 西藏珠穆朗玛峰国家级自然保护区鸟类群落结构与多样性[J]. 生态学报, 33(10): 3056-3064. |
| 王庆忠, 王大科. 1995. 青州仰天山地区夏季鸟类垂直分布的研究[J]. 生态学杂志, 14(1): 33-36. |
| 吴永杰, 杨奇森, 夏霖, 等. 2012. 贡嘎山东坡非飞行小型兽类物种多样性的垂直分布格局[J]. 生态学报, 32(14): 4318-4328. |
| 吴永杰, 雷富民. 2013. 物种丰富度垂直分布格局及影响机制[J]. 动物学杂志, 48(5): 797-807. |
| 吴征镒, 朱彦丞. 1987. 云南植被[M]. 北京: 科学出版杜. |
| 乌日罕, 杨贵生, 魏炜. 2014. 内蒙古阿尔山市北部鸟类区系组成及群落结构[J]. 动物学杂志, 49(1): 94-102. |
| 夏万才, 黎大勇, 范元英, 等. 2014. 白马雪山自然保护区响古箐地区鸟类区系与资源的初步调查[J]. 西华师范大学学报: 自然科学版, 35(4): 339-349. |
| 闫慧, 李敏, 杨贵生. 2011. 内蒙古白银库伦鸟类多样性研究[J]. 四川动物, 30(3): 424-428. |
| 杨岚, 文贤继, 韩联宪, 等. 1994. 云南鸟类志(上卷)非雀形目[M]. 昆明: 云南科技出版社. |
| 杨岚, 杨晓君, 文贤继, 等. 2004. 云南鸟类志(下卷)雀形目[M]. 昆明: 云南科技出版社. |
| 约翰·马敬能, 卡伦·菲利普斯, 何芬奇. 2000. 中国鸟类野外手册[M]. 长沙: 湖南教育出版社. |
| 云南省林业厅. 2003. 白马雪山国家级自然保护区[M]. 昆明: 云南民族出版社. |
| 张荣祖. 1999. 中国动物地理[M]. 北京: 科学出版社. |
| 郑光美. 2011. 中国鸟类分类与名录分布(第二版)[M]. 北京: 科学出版社. |
| 郑作新, 谭耀匡, 梁中宇, 等. 1963. 四川峨眉山鸟类及其垂直分布的研究[J]. 动物学报, 15(2): 317-335. |
| Bhatt D, Joshi KK. 2011. Bird assemblages in natural and urbanized habitats along elevational gradient in Nainital district (western Himalaya) of Uttarakhand State, India[J]. Current Zoology, 57: 318-329. |
| Brehm G, Colwell RK, Kluge J. 2007. The role of environment and mid-domain effect on moth species richness along atropical elevational gradient[J]. Global Ecology and Biogeography, 16(2): 205-219. |
| Colwell RK, Lees DC. 2000. The mid-doman effect: geometric constraints on the geography of species richness[J]. Trends in Ecology and Evolution, 15(2): 70-76. |
| Fu CZ, Hua X, Li J, et al. 2006. Elevational patterns of frog species richness and endemic richness in the Hengduan Mountains, China: geometric constraints, area and climate effects[J]. Ecography, 29(6): 919-927. |
| Mǒrtberg UM. 2001. Resident bird species in urban forest remnants, landscape and habitat perspectives[J]. Landscape Ecology, 16(3): 193-203. |
| Rahdek C. 1995. The elevational gradient of species richness:a uniform pattern[J]. Ecography, 18(2): 200-205. |
| Wu YJ, Colwell RK, Rahbek C, et al. 2013. Explaining the species richness of birds along a subtropical elevational gradient in the Hengduan Mountains[J]. Journal of Biogeography, 40(12): 2310-2323. |
| Wu YJ, Colwell RK, Han NJ, et al. 2014. Understanding historical and contemporary patterns of species richness of babblers along a 5000 m subtropical elevational gradient[J]. Global Ecology and Biogeography, 23(11): 1167-1176. |