 2015, Vol. 17
2015, Vol. 17



2. 陕西省农药管理检定所, 西安 710003
2. Institute for the Control of Agrochemicals, Xi'an 710003, Shaanxi Province, China
杀虫剂被施用后必须进入昆虫体内到达作用部位才能发挥毒效作用。一般而言,杀虫剂可以从昆虫的口腔、表皮及气门3条途径进入昆虫体内,药剂进入后,特别是进入血液后随血液循环分配到各组织器官,会被各种酶系代谢。代谢过程首先是在酶系作用下经历氧化、水解及其他酶促反应生成强极性最终产物,然后再在酶系作用下生成水溶性共轭物排出体外[1, 2]。代谢大多是解毒过程,即将杀虫剂转变成低毒或无毒的产物,也有少数代谢是先转变成毒性更强的化合物(活化),然后再进行解毒代谢。因此,研究药剂在昆虫体内的穿透及代谢,对于研究药剂的作用方式及作用机制、采用科学的制剂加工及施药技术都有重要意义。
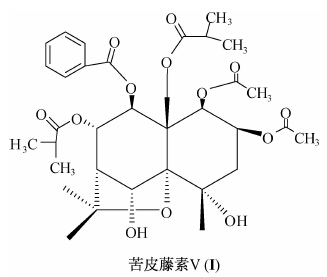
杀虫植物苦皮藤Celastrus angnlatus的杀虫活性成分是以苦皮藤素V(结构式见I)为代表的一系列二氢沉香呋喃多元醇酯类化合物[3, 4, 5, 6]。以苦皮藤根皮提取物为原料,已成功研制了0.2%苦皮藤素乳油[7]、0.15%苦皮藤素微乳剂[8]及0.2%苦皮藤素水乳剂,其中0.2%苦皮藤素水乳剂已于2013年登记。已有的研究结果表明,苦皮藤素Ⅴ无触杀和熏蒸作用,只有胃毒作用,即只有药剂被摄入昆虫消化系统后,试虫才会表现出中毒症状,该过程约需2 h[9, 10]。药剂对昆虫没有触杀作用是因为药剂不能穿过昆虫表皮进入体内,还是能穿过表皮但在体内传导过程中不能到达靶标部位呢?已有的研究结果表明,苦皮藤素Ⅴ的胃毒作用在小地老虎与粘虫幼虫之间有明显的选择毒杀作用:在胃毒剂量为37.59 μg/g时,4龄粘虫幼虫已表现出明显的中毒症状,24 h死亡率可达80%以上,而在胃毒剂量高达10 500 μg/g时,4龄小地老虎幼虫仍无中毒症状,24 h无一虫死亡[11, 12]。目前关于苦皮藤素对昆虫的选择毒性机理方面研究,仅见昆虫中肠pH值及解毒磷酸酯酶活性差异的报道[11, 12]。为此,笔者采用高效液相色谱法研究了苦皮藤素Ⅴ在粘虫和小地老虎幼虫体内的穿透及代谢差异,以进一步探讨苦皮藤素V对昆虫毒性的选择性机理。
苦皮藤素Ⅴ(celangulin V)标准品,含量98%,西北农林科技大学农药研究所提供。乙腈,色谱纯,美国Tedia公司;甲醇、丙酮和二甲亚砜,分析纯,天津博迪化工股份有限公司。Finnegan Suveyor高效液相色谱仪,具二极管阵列检测器,美国Thermo公司;Spherrisorb C18柱(250 mm×4.6 mm,5 μm),美国Waters公司;C18固相萃取柱(500 mg,6 mL),美国Agilent公司。
1.1.2 供试试虫东方粘虫Mythimna separata 6龄幼虫(蜕皮后第2天),小地老虎Agrotis ypsilon 5龄幼虫(蜕皮后第2天),均由西北农林科技大学农药研究所养虫室提供。 1.2 苦皮藤素Ⅴ在昆虫体内的穿透能力 1.2.1 从体壁到血腔的穿透
将试虫分放于24孔板中,饥饿8 h 后在其前胸背板点滴1 μL 40 mg/mL 的苦皮藤素V丙酮溶液,对照点滴1 μL 丙酮。分别于点滴处理后1、2、4、6、8及16 h各取1组(5头)试虫,刺破腹足收集血淋巴于2.0 mL离心管中,-20 ℃冰箱保存备用。每处理重复4次。
1.2.2 从中肠到血腔的穿透将试虫分放于24孔板中饥饿24 h 后,以载药量为20 μg/片的小麦或油菜叶片(4 mm×5 mm)饲喂,每虫1片。标记每头试虫取食完叶碟的时间,分别于取食后1、2、4、6、8及16 h各取1组(5头)试虫,刺破腹足收集血淋巴于2.0 mL离心管中,-20 ℃冰箱保存备用。每处理重复4次。
1.2.3 从血腔到中肠的穿透将试虫饥饿8 h后,以乙醚麻醉,注射0.4 μL 50 mg/mL的苦皮藤素V二甲亚砜溶液,对照注射0.4 μL二甲亚砜。分别于处理后1、2、4、6、8及16 h各取1组(10头)试虫,以试虫头部在2 mL离心管口剐蹭,收集试虫返吐液,于-20 ℃冰箱中保存备用。每处理重复4次。
1.2.4 血淋巴及返吐液中苦皮藤素Ⅴ含量分析 1.2.4.1 样品前处理在100 μL粘虫血液或返吐液中添加0.5 μg苦皮藤素V标准品,混匀后加入400 μL甲醇,离心(3 000 r/min)后取上清液300 μL,加入2 mL重蒸水稀释后通过预先活化的固相萃取柱,分别以V(甲醇)∶V(水)=0∶100,20∶80,50∶50,80∶20及100∶0 淋洗,各收集2 mL洗脱液。采用高效液相色谱仪(HPLC)分析各洗脱液中苦皮藤素V的含量,计算洗脱率,从而确定最佳淋洗液。
在C18固相萃取柱中,先用2 mL甲醇活化,再用2 mL重蒸水淋洗。重复1次,备用。取试虫血淋巴或返吐液100 μL于2 mL离心管中,加入400 μL甲醇,摇匀后经3 000 r/min离心10 min。取上清液300 μL于5 mL离心管中,加入2 mL重蒸水稀释,过固相萃取柱,弃去淋出液。先以2.0 mL V(甲醇)∶V(水)=1∶1溶液淋洗,弃去洗脱液,再以2.0 mL甲醇淋洗,收集洗脱液,待HPLC检测。
1.2.4.2 HPLC检测条件流动相V(甲醇)∶V(乙腈)∶V(水)=40∶30∶30;流速1.0 mL/min;检测波长230 nm;进样量10 μL。
1.2.4.3 标准曲线的建立以甲醇为溶剂配制质量浓度分别为2.25、1.125、0.56、0.28、0.14及0.07 μg/mL的苦皮藤素V系列标准溶液,按所建立的色谱条件进样,每浓度重复3次。根据不同进样量及对应的峰面积绘制苦皮藤素V的标准曲线,计算线性方程及相关系数。
1.2.4.4 分析方法的准确度及精密度分别在粘虫和小地老虎血淋巴、返吐液及幼虫匀浆液中添加2.0 和4.0 μg/mL苦皮藤素V标准溶液,按本研究方法进行样品前处理及色谱分析,计算添加回收率及相对标准偏差,以评价方法的准确度和精确度。试验重复5次。
1.3 苦皮藤素V在昆虫体内的代谢能力测定 1.3.1 试虫处理将试虫于24孔板中饥饿24 h后,以每片含有20 μg苦皮藤素V的小麦或油菜叶片(4 mm×5 mm)饲喂,每虫1片,标记每头试虫取食完叶碟的时间,分别于取食后0、1、2、4、8、16、24及36 h各取1组试虫(5头)及其排出粪便,放入匀浆管内,加入5 mL甲醇,电动匀浆2 min。将匀浆液转入25 mL容量瓶中,用甲醇洗涤转头及匀浆管3次,每次5 mL,一并转入容量瓶并定容至25 mL。取2.0 mL匀浆液于离心管内,在3 000 r/min下离心10 min,取上清液按1.2.4节方法进行前处理。试验重复3次。
1.3.2 苦皮藤素V含量分析色谱分析条件同1.2.4.2节。 2 结果与分析 2.1 苦皮藤素V分析方法的建立
线性关系研究结果表明,在0.07~2.25 μg/mL范围内,苦皮藤素V进样质量浓度与峰面积呈良好的线性关系,其线性方程为y=48 438x+1 560.1,相关系数r为0.999 8。准确度和精确度测定结果见表 1。以苦皮藤素V的峰面积信噪比≥3为检出限,得苦皮藤素V在6种供试样品中的检出限为0.5 ng,最低检测浓度为0.05 μg/mL。表明所建立的HPLC方法可以满足本研究中苦皮藤素V含量分析的要求。
| 表 1 苦皮藤素Ⅴ在昆虫血淋巴、返吐液及虫体匀浆液中的添加回收率及相对标准偏差(n=5) Table 1 Recoveries of celangulin V in the haemolymph,regurgitation juice and homogenates of the tested larvae(n=5) |
样品前处理测定结果表明:以V(甲醇)∶V(水)=0∶100,20∶80和50∶50洗脱柱床,洗脱液均未检出苦皮藤素V;用V(甲醇)∶V(水)=80∶20洗脱,苦皮藤素V的洗脱率为70.91%;用甲醇洗脱,洗脱率为99.94%。最终选用以甲醇为淋洗液。
2.2 苦皮藤素Ⅴ在昆虫体内的穿透作用2.2.1 从体壁到血腔的穿透
在所设定的HPLC条件下,以40 μg/头的剂量在6龄粘虫幼虫或5龄小地老虎幼虫前胸背板进行苦皮藤素V点滴处理,分别于1、2、4、6、8及16 h取血淋巴,均未检测到苦皮藤素V。
2.2.2 从中肠到血腔的穿透
粘虫及小地老虎幼虫经胃毒处理不同时间后血淋巴中苦皮藤素V的含量见表 2。
| 表 2 胃毒处理粘虫和小地老虎血淋巴中苦皮藤素V的含量(n=4)
Table 2 Contents of celangulin V in hemolymph of M. separata and A.ypsilon by stomach poison(n=4)
|
从表 2可以看出:苦皮藤素V被摄入消化道后很快穿过肠壁细胞进入血腔,1 h后粘虫和小地老虎血淋巴中苦皮藤素V的含量分别达1.74 和 1.65 μg/mL,4 h分别达峰值(4.54和 5.81 μg/mL),此后由于代谢作用含量下降,16 h后分别降至0.92和0.98 μg/mL。 2.2.3 从血腔到中肠的穿透
在所选HPLC条件下,以20 μg/头的剂量对6龄粘虫幼虫或5龄小地老虎幼虫进行苦皮藤素V注射处理,分别于1、2、4、6、8及16 h取返吐液,均未检测到苦皮藤素V。
2.3 苦皮藤素Ⅴ在昆虫体内的代谢
粘虫和小地老虎幼虫摄食苦皮藤素V后,体内苦皮藤素V的含量见表 3,其典型色谱图见图 1。由表 3可以看出,苦皮藤素V随食物摄入试虫体内后很快被降解。1 h后其在粘虫和小地老虎幼虫体内的降解率分别为10.85%和25.46%,24 h后分别为71.13%和96.63%。苦皮藤素V在粘虫体内的代谢方程为ct=92.3e-0.052 8t,R2=0.923 5,半衰期为13.1 h,43.6 h后可代谢90%;在小地老虎体内的代谢方程为ct=88.3e-0.126 8t,R2=0.958 4,半衰期为5.5 h,18.2 h后可代谢90%。
| 表 3 苦皮藤素V在粘虫和小地老虎幼虫体内的代谢率(n=4) Table 3 The metabolism rates of celangulin V in the larvae of M. separata and A.ypsilon(n=4) |
|
a、b、c:小地老虎0、8和24 h;d、e、f:粘虫0、8和24 h。 a,b,c: A.ypsilon at 0,8 and 24 h;d,e,f: M. separata at 0,8 and 24 h. 图 1 苦皮藤素V在粘虫和小地老虎幼虫体内的代谢色谱图 Fig. 1 Chromatogram of metabolisms of celangulin V in the larvae of M. separata and A.ypsilon |
本研究结果表明,通过昆虫局部点滴处理,苦皮藤素V难以穿透试虫表皮进入血腔,并难以从血腔进入中肠,但通过载毒叶片饲喂处理,其很容易穿透肠壁进入血腔。其中难以穿透表皮的原因除了化合物本身的分子属性外,还与昆虫体壁结构有关。昆虫体壁有多层保护结构,如上表皮的护蜡层、蜡层及角质精层,还有经硬化的外表皮,这些都可以阻止药物穿过昆虫体壁进入血腔。此外,局部点滴处理本身亦不利于药剂穿透:当苦皮藤素V的丙酮溶液点滴到昆虫表皮后,丙酮很快挥发,致使化合物以固态形式附着于小面积的表皮上(不排除试虫在爬行过程中有失落的可能),因此难以穿透。至于苦皮藤素V容易穿透肠壁进入血腔而难以从血腔穿透肠壁进入中肠的原因,可能涉及药物载体等问题,有待进一步研究。
研究药物在靶标生物中的穿透行为可能为药物作用机理研究提供有益启示。由于苦皮藤素V不能穿透昆虫表皮进入体内,因此没有触杀活性。苦皮藤素V很容易从中肠进入血腔,然而虽然昆虫神经系统、呼吸系统浸泡在血淋巴中,但苦皮藤素V并未表现出神经毒剂或呼吸毒剂的作用症状。采用血腔注射的方式将苦皮藤素V注入血腔(剂量为6龄粘虫40 μg/头),发现苦皮藤素V并不能穿透肠壁到达中肠,试虫亦未表现出任何中毒症状,这说明苦皮藤素V的作用靶标不在神经系统或呼吸系统。迄今的研究表明,苦皮藤素V惟有被摄入消化系统达到中肠才会引起试虫中毒死亡,而且其症状特殊:上吐下泻,摄毒约2 h后试虫因大量失去体液(水分和血淋巴)而死亡,死虫虫体显著缩短。苦皮藤素V在虫体的这种穿透行为,提示其作用部位在试虫消化系统,作用靶标可能位于中肠肠壁细胞。
关于杀虫剂的选择性,按Ripper等的说法可分为生态选择性(ecological selectivity)和生理选择性(physiological selectivity)[13]。生态选择性是在毒物存在环境中,一种生物中毒死亡,另一种则可能以某种方式避免接触毒物而存活的现象,这是一种外在的选择性,其原因主要是各种生物行为习性的差异;生理选择性是两种生物都暴露在有毒环境下,但其中一种由于某些生理生化上的机制,有更高的耐受力而存活的现象,这是一种内在的选择性。Hollingworth等认为杀虫剂选择性的生理生化机制主要有5个方面:穿透差异(外阻隔层穿透差异、内阻隔层穿透差异、非靶标部位结合差异)、排毒效能差异、代谢差异(解毒代谢差异、活化代谢差异)、作用部位差异(靶标部位敏感性差异、独特的作用靶标)及综合作用[14, 15]。本研究表明,在粘虫和小地老虎幼虫中,苦皮藤素V均不能从体壁穿透到血腔或从血腔穿透到中肠,但很容易从中肠穿透到血腔,而且穿透速率无差异,说明苦皮藤素V的选择性与穿透能力无关。苦皮藤素V在小地老虎幼虫体内的代谢解毒速率远大于其在粘虫幼虫体内的,其半衰期分别为5.5和13.1 h,说明苦皮藤素V的选择作用和其代谢解毒有一定关系。一般来说,由于代谢差异造成的药剂选择作用往往表现为对试虫毒力的差异,即对代谢快的试虫毒力低,对代谢慢的试虫毒力高。苦皮藤素V对小地老虎幼虫的投毒剂量即使是粘虫幼虫的279倍亦不表现中毒症状,说明代谢差异不应是其主要的选择机制,靶标敏感性差异或特异性靶标可能才是根本的选择机制。由于苦皮藤素V具体的作用靶标尚不确定,因此,其选择作用机制是否基于靶标敏感性差异或具有靶标特异性尚需深入研究。
| [1] | Wilkinson C F. Insecticide Biochemistry and Physiology[M]. London: Heyden, 1976. |
| [2] | 吴文君. 农药学原理[M]. 北京: 中国农业出版社, 2000. Wu Wenjun. The Principle of Pesticide Science[M]. Beijing: Chinese Agricultural Press, 2000.(in Chinese) |
| [3] | 吴文君, 姬志勤, 胡兆农, 等. 杀虫植物苦皮藤中的有效成分及其生物活性[J]. 华中师范大学学报: 自然科学版, 2005, 39(1): 50-53. Wu Wenjun, Ji Zhiqin, Hu Zhaonong, et al. Advances in research on insecticidal plant Celastrus angulatus: chemistry of the active components[J]. J Central Chin Normal Univ: Nat Sci, 2005, 39(1): 50-53.(in Chinese) |
| [4] | 吴文君. 从天然产物到新农药创制: 原理·方法[M]. 北京: 化学工业出版社, 2006: 242-291. Wu Wenjun. From Natural Products to the Development of New Pesticides: Principles & Methods[M]. Beijing: Chemical Industry Press, 2006: 242-291.(in Chinese) |
| [5] | 吴文君. 植物杀虫剂苦皮藤素研究与应用[M]. 北京: 化学工业出版社, 2011. Wu Wenjun. Research and Application of Botanical Insecticides Celangulins[M]. Beijing: Chemical Industry Press, 2011.(in Chinese) |
| [6] | 吴文君, 刘惠霞, 胡兆农, 等. 从天然产物到新农药创制——杀虫植物苦皮藤研究进展[J]. 昆虫知识, 2008, 45(6): 845-851. Wu Wenjun, Liu Huixia, Hu Zhaonong, et al. Advances in research of insecticidal plant Celastrus angulatus [J]. Chin Bull Entomol, 2008, 45(6): 845-851.(in Chinese) |
| [7] | 吴文君, 刘惠霞, 姬志勤, 等. 植物杀虫剂0.2%苦皮藤素乳油的研究与开发[J]. 农药, 2001, 40(3): 17-19. Wu Wenjun, Liu Huixia, Ji Zhiqin, et al. Research and development on the botanical insecticide of 0.2% celangulins emulsifiable concentrate[J]. Pesticides, 2001, 40(3): 17-19.(in Chinese) |
| [8] | 刘小凤, 相建业, 胡想顺, 等. 0.15%苦皮藤素微乳剂防治小菜蛾田间药效试验[J]. 农药, 2003, 42(1): 44-45. Liu Xiaofeng, Xiang Jianye, Hu Xiangshun, et al. 0.15% celangulin microemulsion for controlling P.xylostella in field efficacy[J]. Pesticides, 2003, 42(1): 44-45.(in Chinese) |
| [9] | 吴文君, 曹高俊. 植物性杀虫剂苦皮藤作用方式的初步研究[J]. 西北农学院学报, 1982, 10(1): 75-80. Wu Wenjun, Cao Gaojun. Preliminarily study on mode action of Celastrus angulatus[J]. J Northwest Sci-Tech Univ Agric Forest: Nat Sci Ed, 1982, 10(1): 75-80.(in Chinese) |
| [10] | 吴文君, 陈广泉, 王兴林. 苦皮藤提取物对玉米象种群的控制作用及机理[J]. 粮食储藏, 1988, 17(6): 9-14. Wu Wenjun, Chen Guangquan, Wang Xinglin. Insecticidal activity and mechanism of extract of Chinese bittersweet against maize weevil, Sitophilus zeamais Motschulsky[J]. Grain Storage, 1988, 17(6): 9-14.(in Chinese) |
| [11] | 刘惠霞, 吴文君, 姬志勤, 等. 苦皮藤毒杀成分对昆虫的选择毒杀作用及其机制研究[J]. 西北农业学报, 1998, 7(2): 41-44. Liu Huixia, Wu Wenjun, Ji Zhiqin, et al. Studies on the selective toxicity and selective mechanism of poisonous ingredient celangulin V of Celastrus angulatus against the insect pests[J]. Acta Agriculturae Boreali-occidentalis Sinica, 1998, 7(2): 41-44.(in Chinese) |
| [12] | 刘惠霞, 杨从军, 吴昊, 等. 苦皮藤素Ⅴ对昆虫选择毒性机理的进一步研究[J]. 西北农林科技大学学报: 自然科学版, 2002, 30(2): 83-86. Liu Huixia, Yang Congjun, Wu Hao, et al. Further studies on the mechanism of selective toxicity of celangulin V[J]. J Northwest Sci-Tech Univ Agric Forest: Nat Sci Ed, 2002, 30(2): 83-86.(in Chinese) |
| [13] | Ripper W E, Abel A L, Greenslade R M, et al. Pest Control: Farmers' Handbook[M]. Cambridge: Pest Control Limited, 1952. |
| [14] | Hollingworth R M. Chemistry, biological activity, and uses of formamidine pesticides[J]. Environ Health Persp, 1976, 14: 57-69. |
| [15] | 张宗炳. 杀虫药剂的分子毒理学[M]. 北京: 农业出版社, 1987. Zhang Zongbing. Molecular Toxicology Insecticides[M]. Beijing: Agriculture Press, 1987.(in Chinese) |