
文章信息
- 刘伦, 王超, 姚改芳, 王德孚, 吴俊. 2015.
- LIU Lun, WANG Chao, YAO Gaifang, WANG Defu, WU Jun. 2015.
- 外源生长调节剂处理对‘满天红’ב砀山酥梨’后代成花的影响
- Effects of exogenous growth regulator treatment on floral initiation of pear progenies from hybrids'Mantianhong’×'Dangshansuli’
- 南京农业大学学报, 38(3): 381-388
- Journal of Nanjing Agricultural University, 38(3): 381-388.
- http://dx.doi.org/10.7685/j.issn.1000-2030.2015.03.005
-
文章历史
- 收稿日期:2014-08-26




梨属于木本植物,要经历童期、过渡期、成年期之后,才能开花结果。一般来说,果树童期要经过4~8年的时间,在这个阶段,除了改变遗传因素外,任何外部措施都不能诱导其开花。但如果在过渡期利用外部条件进行诱导,可以加速其成花。研究表面已通过童期的3年生实生苹果树喷施生长调节剂处理后可以成花,最低成花节位为77节,喷施2次50 mg·kg-16-BA+1 000 mg·kg-1乙烯利可明显加快苹果实生树成花[1]。Chacko等[2]报道利用乙烯利可以诱导芒果实生苗提早开花结果。有研究表明,细胞分裂素有促花机制,人们常将细胞分裂素作为根生长状态的信号因子,并将它同果树的负载量联系在一起[3]。目前,在果树上有通过喷施外源生长调节剂使实生苗开花结果的研究报道,但是大都集中在对成花表型的观察,包括始花节位数的观察,枝类的构成,激素的选择和喷施时间等方面,或者只研究外源生长调节剂对内源激素的影响等[4, 5],并没有深入研究其内在的作用机制。对于果树育种工作者来说,由于果实相关的性状评价必须要等到结果后才能开展,因此,寻找促使实生苗提早开花结果的办法,有利于缩短育种周期,加速育种进程,对于果树育种工作具有十分重要的意义。目前在梨上尚未开展促进杂交后代成花转变的研究报道,因此,本研究以5年生的‘满天红’ב砀山酥梨’杂交后代未开花的实生苗为材料,通过喷施乙烯利(ethephon,Eth)、6-BA以及二者的混合液,研究外源生长调节剂处理对成花的影响,并通过对成花相关基因表达、内源激素含量,以及营养物质含量变化等研究,初步探索促进成花的相关因素及其作用机制,为促进梨杂种实生后代提早开花、缩短育种周期提供理论依据。 1 材料与方法 1.1 试验材料和处理
选取2008年定植且2012年仍未开花的长势一致的‘满天红’ב砀山酥梨’为试验材料,分别于花芽分化期(2012年6月25日和7月3日)连续2次对实生树进行4个喷施处理:Ⅰ)100 mg·kg-16-BA;Ⅱ)1 000 mg·kg-1Eth;Ⅲ)100 mg·kg-16-BA+1 000 mg·kg-1Eth;CK)清水对照。喷施程度以全树的叶片滴水为止。单株小区,每个处理3次重复。分别于2012年5月20日(花芽分化前)、7月10日(花芽分化期)和9月24日(花芽分化末期)随机采集4个处理树体的芽用液氮处理后,-80 ℃超低温冰箱中保存。于2013年3月19日分别统计4个处理每个单株形成的花芽数量。 1.2 芽RNA提取及cDNA第一链的合成
采用改良的CTAB法[6]提取梨花芽组织RNA。采用10g·L-1的琼脂糖凝胶电泳检测RNA质量,用微量核酸分析仪NanoDrop(Thermo)测RNA的浓度。消化后的RNA经65 ℃ 5 min变性后置于冰上。配制反应液为:前一个步骤中变性的RNA 11 μL,5×RT Buffer 2 μL,RT Enzyme Mix 0.5 μL,Primer Mix 0.5 μL,合成体系的总体积为20 μL。程序为:37 ℃ 15 min,98 ℃ 5 min,4 ℃ 5 min,于-20 ℃保存备用。
反转录结束后利用NanoDrop检测合成cDNA的浓度,稀释到100 ng·μL-1用于实时定量分析。 1.3 实时荧光定量PCR反应
利用实时荧光定量PCR方法,以梨EFα1(AY338250)[7]基因为内参基因,检测成花相关基因LEAFY(LFY)、TERMINAL FLOWER1(TFL1)和FLOWERING LOCUS T(FT)的时空表达特性。以已克隆的梨LFY、TFL1、FT和EFα1序列为模板,利用PrimerPremier5.0设计荧光定量引物,引物序列见表 1。
| 基因 Gene | PCR产物长度/bp PCR product length | 引物序列(5′→3′) Primer sequence | 用途 Usage |
| LFY | 226 | F/R:AGGGAGCACCCGTTCATCGT/GCCGCATCTTTGGCTTGTTG | RT-PCR |
| TFL1 | 144 | F/R:GCCGCATCTTTGGCTTGTTG/GCCGCATCTTTGGCTTGTTG | RT-PCR |
| FT | 220 | F/R:AGCCCAAGTGACCCCAACCT/CGGCGAAGTCTCTGGTATTGAAG | RT-PCR |
| EFα1 | 165 | F/R:GGTGTGAAGCAGATGATTT/TCACCCTCAAACCCAGATAT | RT-PCR |
PCR反应条件为:95 ℃ 5 min;94 ℃ 10 s,60 ℃ 30 s,72 ℃ 30 s,共40个循环;72 ℃ 3 min。所有样品均重复3次,采用Light Cycler 480荧光定量软件进行数据采集,用MicrosoFT 2003进行数据分析。用2-ΔΔCT计算基因的表达量。CT值表示实时定量PCR中荧光信号达到所设定的阈值时,所经历的反应循环数。
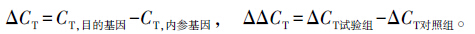
芽内植物激素的提取参考曹尚银等[8]的方法。从每个处理的3棵树上采的芽混合研磨后提取內源激素。采用高效液相色谱法,液相色谱仪为Waters1525系统。色谱条件为hypersil ODS C18柱,流动相:甲醇和超纯水,加入0.6%(体积分数)的冰乙酸,流速0.7 mL·min-1,进样量:5 μL。检测器类型是Waters 2487紫外检测器。用外标定量法测定GA3、ZT、ABA、IAA的含量,每个处理的样品测定3次。标样IAA、GA3、ABA、ZT和色谱甲醇均购自Sigma公司。 1.5 芽内营养物质含量的测定
采用蒽酮比色法[9]测定芽内可溶性糖含量和淀粉含量,用凯式定氮法[10]测定总氮含量。每个处理3棵树上采集的芽混合研磨后提取芽内营养物质。每个处理的样品测定3次。根据可溶性糖、淀粉和总氮的含量,计算出C/N值。 1.6 数据分析 利用SPSS 19.0软件对试验数据进行方差分析和相关性分析。 2 结果与分析 2.1 生长调节剂处理对杂交后代花芽数量的影响
由图 1可知:不同生长调节剂处理显著增加了单株成花数,其中6-BA处理和6-BA+Eth混合处理的效果要优于Eth处理。喷施6-BA和Eth的混合液形成的花芽数量最多,效果最佳。说明植物生长调节剂处理可以促进成花转变,加速树体由童期向成年期的过渡。
 |
图 1 生长调节剂对花芽形成数的影响 Fig. 1 The effect of growth regulator on the amount of floral buds
1)Ⅰ.100 mg·kg-16-BA;Ⅱ.1 000 mg·kg-1Eth;Ⅲ.100 mg·kg-16-BA+1 000 mg·kg-1Eth;CK.对照Control 2)不同字母表示不同处理之间差异显著(P <0.05)。The different letters indicate significant difference at 0.05 level among different treatments.The same as follows. |
由图 2可见:生长调节剂处理前(5月20日)不同处理和对照的ZT(玉米素)和ABA含量均较低且差异不显著。随着花芽分化的进行,ZT和ABA含量开始增加,且在7月花芽开始分化期含量最高,之后下降。各处理的ZT含量在花芽分化期和花芽分化末期显著高于对照组。各处理的ABA的含量只在花芽分化期显著高于对照。可见,喷施外源生长调节剂处理有利于提高ZT和ABA的含量。不同处理和对照在花芽未分化时GA3和IAA含量较高且无显著差异。随着花芽分化的进行,GA3和IAA含量均下降,并且都在花芽分化接近结束(9月份)时达到最低。在花芽分化期和花芽分化末期,各处理的GA3和IAA含量显著低于对照。可见,喷施外源生长调节剂后有利于降低GA3和IAA含量。
 | 图 2 生长调节剂对芽内源激素含量的影响 Fig. 2 The effect of growth regulator on endogenous hormone content in buds |
由图 3-A、B可见:在花芽分化前,促进类激素ZT、ABA与抑制类激素GA3和IAA的比值均在花芽未分化时保持低水平;随着花芽分化开始,比值升高,并且各处理均显著高于对照。不同处理的ZT/GA3和ZT/IAA的比值在花芽开始分化时骤升,之后有所下降,但是依然保持高水平。对照在各时期差异不显著。可见,喷施生长调节剂有利于提高ZT/GA3和ZT/IAA的值。ZT/IAA各时期不同处理间差异不显著。
 | 图 3 生长调节剂处理对芽内源激素比值的影响Fig. 3 The effect of growth regulator on the ratio of endogenous hormone in buds | |
花芽分化前ABA/IAA和ABA/GA3比值处于较低水平,且处理与对照之间没有显著差异(图 3-C、D)。随着花芽分化的进行ABA/IAA和ABA/GA3比值明显升高,处理显著高于对照,每个时期不同处理之间差异不显著。表明喷施生长调节剂后ABA/IAA和ABA/GA3比值明显增加。 2.3 生长调节剂对花芽营养物质的影响
由图 4可见:花芽分化前,各处理和对照芽内可溶性糖和C/N值含量处于较低水平,且无显著差异;在花芽分化期和分化末期,3个处理芽内可溶性糖含量较分化前显著升高,对照则与分化前无明显变化,且显著低于处理组,但3个处理之间无显著差异。表面生长调节剂处理促进了芽内可溶性糖的积累,有利于提高C/N值。
 | 图 4 生长调节剂处理对芽营养物质含量的的影响Fig. 4 The effect of growth regulator on nutrients content in buds | |
花芽分化前,各处理和对照芽内淀粉和氮含量处于较高水平,且无有显著差异;在花芽分化期和分化末期,3个处理的芽内淀粉含量和氮含量较分化前显著降低,对照则无明显变化,且显著高于处理。淀粉含量在处理之间差异不显著。表面生长调节剂处理使淀粉和氮含量降低。 2.4 生长调节剂对成花相关基因LFY、TFL1和FT表达的影响
由图 5可见:在生长调节剂喷施之前,不同处理和对照之间LFY和FT基因的表达量较低,且无明显差异;但是在花芽分化期和分化末期时,喷施生长调节剂的3个处理中LFY和FT基因的表达量均显著高于对照,且在花芽分化期到达最高,在分化末期时表达量下降。处理Ⅲ的LFY基因的表达量增加最多,其次是处理Ⅰ,处理Ⅱ相对低;处理ⅠFT基因的表达量最高,处理Ⅰ和处理Ⅱ之间差异不显著。表面不同生长调节剂处理对LFY基因表达的促进效果与成花数量的促进效果相一致。
 | 图 5 生长调节剂处理对成花相关基因LFY(A)、TFL 1 (B)和FT(C)表达的影响Fig. 5 The effect of growth regulator on the expression of flowering-related genes LFY(A),TFL 1 (B) and FT | |
不同处理和对照TFL1基因的表达量在花芽未分化时最高,随着花芽分化的进行TFL1的表达量呈现下调的趋势,且各处理均低于对照,至分化末期时,TFL1基因的表达量极低。各处理的TFL1基因表达量下降速度高于对照。在花芽分化期,处理Ⅱ的TFL1的表达量稍高于处理Ⅰ和处理Ⅲ,与此对应,处理Ⅱ最终形成的花芽个数也偏低,表明TFL1基因的表达抑制了花芽的分化。 2.5 生长调节剂对各成花因素之间相关性的影响
为明确不同成花因素以及基因表达之间的关系,根据促进成花转变效果最好的处理Ⅲ(喷施 100 mg·kg-16-BA+1 000 mg·kg-1Eth)测定的各项数据,对各因素之间的相关性进行分析(表 2)。
| LFY | TFL1 | FT | ZT | GA3 | IAA | ABA | ZT/GA3 | ZT/IAA | ABA/GA3 | ABA/IAA | 可溶性糖 Soluble sugar | 淀粉 Starch | 氮 Nitrogen | C/N |
| LFY | 1 | ||||||||||||||
| TFL1 | -0.847** | 1 | |||||||||||||
| FT | 0.839** | -0.457 | 1 | ||||||||||||
| ZT | 0.933** | -0.633 | 0.952** | 1 | |||||||||||
| GA3 | -0.759* | 0.968** | -0.310 | -0.531 | 1 | ||||||||||
| IAA | -0.845** | 0.966** | -0.492 | -0.627 | 0.923** | 1 | |||||||||
| ABA | 0.959** | -0.821** | 0.860** | 0.904** | -0.687* | -0.825** | 1 | ||||||||
| ZT/GA3 | 0.961** | -0.895** | 0.726* | 0.883** | -0.861** | -0.853** | 0.901** | 1 | |||||||
| ZT/IAA | 0.970** | -0.915** | 0.751* | 0.855** | -0.837** | -0.939** | 0.952** | 0.949** | 1 | ||||||
| ABA/GA3 | 0.902** | -0.985** | 0.563 | 0.729* | -0.951** | -0.949** | 0.875** | 0.951** | 0.948** | 1 | |||||
| ABA/IAA | 0.884** | -0.955** | 0.590 | 0.695* | -0.877** | -0.988** | 0.895** | 0.868** | 0.964** | 0.950** | 1 | ||||
| 可溶性糖 Soluble sugar | 0.991** | -0.874** | 0.818** | 0.906** | -0.777* | -0.869** | 0.981** | 0.956** | 0.977** | 0.927** | 0.914** | 1 | |||
| 淀粉 Starch | 0.862** | -0.977** | 0.533 | 0.660 | -0.920** | -0.992** | 0.864** | 0.874** | 0.948** | 0.966** | 0.992** | 0.893** | 1 | ||
| 氮 Nitrogen | -0.895** | 0.965** | -0.520 | -0.708* | 0.956** | 0.930** | -0.822** | -0.942** | -0.923** | -0.979** | -0.914** | -0.906** | -0.931** | 1 | |
| C/N | 0.943** | -0.936** | 0.624 | 0.792* | -0.911** | -0.900** | 0.868** | 0.970** | 0.941** | 0.969** | 0.899** | 0.945** | 0.906** | -0.989** | 1 |
由表 2可知:不同基因之间,LFY与TFL1呈极显著负相关;LFY与FT呈极显著正相关;FT与TFL1呈负相关,但是相关性不显著,说明成花相关基因LFY、FT和TFL1存在着此消彼长的作用趋势。不同激素之间,ZT与GA3呈负相关,但是相关性不显著,与IAA呈显著负相关,与ABA呈极显著正相关;GA3与IAA显著正相关,与ABA显著负相关;IAA与ABA极显著负相关,说明内源激素之间有相互促进和相互制约的关系。ZT和ABA在花芽分化过程中促进分化,GA3和IAA在花芽分化过程中抑制分化,同类激素之间有相互促进的作用,异类激素有相互抑制的作用。可溶性糖与淀粉含量、氮含量有极显著相关性,淀粉含量和氮含量呈极显著正相关。此外,从表 2也可以看出,基因、内源激素和营养物质之间也有较大相关性,说明这3方面的因素之间也相互影响,进一步说明成花是一个复杂的过程,需要多因素共同参与调控。 3 讨论
梨的杂交后代播种发芽后,幼苗处在童期,即无开花能力时期。前人对木本果树由童期向成年期过渡的生理机制进行了大量研究,其中多因子假说[11]逐渐得到学术界的认同。该假说认为,花诱导过程中,多种激素、代谢物、营养等化学物质都有参与,其中碳水化合物在这个过程中起到了很重要的作用。本研究表明:外源植物生长调节剂处理,影响了处于成花转变时期幼苗的内源激素、营养物质和基因表达水平,它们与成花均表现一定的相关性,从而进一步说明了成花的复杂性。
为了缩短童期到成年期的时间,前人对诱导果树成花的技术进行了试验研究。过国南等[5]通过对3年生苹果杂交实生苗连续喷施生长延缓剂,发现多效唑能有效抑制苹果杂交苗的营养生长,促使杂交苗早开花。多效唑与适宜浓度的乙烯利混合使用,有利于提高首次开花株率,增加单株平均花序数。已有研究表明,梨树发育过程中,在生理分化期喷施6-BA抑制花芽分化,在形态分化期喷施则促进花芽分化[5];而在荔枝[12]、梅[13]和苹果[14]的研究中发现,6-BA也表现为促进花芽分化的作用。此外,对处于营养生长旺盛的‘苹果梨’喷施乙烯利有利于花芽分化[15]。本研究中选择的梨实生树为5年生个体,正处于童期到成年期的转变期,喷施6-BA、乙烯利以及两者混合液均促进了幼苗的花芽分化,其中喷施乙烯利和6-BA的混合液效果最好,其次为6-BA处理。可见,6-BA和乙烯利都有促进花芽分化的作用。
有研究报道,乙烯利处理后,IAA含量均显著下降,ABA含量显著升高,GA4/ABA比值显著减小,并且各试材GA4/ABA的比值变化与其节间长度变化较吻合[16]。花芽生理分化期内高水平的ABA含量不能促进花芽分化[17],在生理分化期花芽中ABA含量保持低水平,而在花芽分化期其含量明显高于对照。ABA通过增加液泡中的糖含量,进而提高植株吸收营养物质的能力,来达到促进成花的作用[18]。另外,在花芽转变过程中花芽内源激素之间也相互影响。同为促进花芽分化的激素之间有相互促进的作用。同为抑制花芽分化的激素之间有相互抑制的作用。
碳水化合物对苹果花芽启动的影响较小,但是对花芽分化的质量起着重要作用[19]。本研究中,生长调节剂处理后在花芽分化期间可溶性糖和淀粉有较大程度的积累,同时花芽分化时保持高水平的C/N,都有利于促进成花。这与郭金丽等[20]在苹果上的研究结果一致。
LFY作为调控开花的一个重要转录因子,对促进植物开花有重要作用。转入拟南芥中促进成花启动的LFY基因的甜橙和枳橙比未转化的植株提早3~5年开花,童期明显缩短[21]。在梨中,PbLFY基因不仅在营养组织中表达,也在花器官中表达,与梨的花芽分化与开花关系密切[22]。已有报道表明,杨树PdFT基因和三叶桔CiFT基因的组成型表达,可以缩短幼龄树的童期,促进成年植株开花[23, 24]。研究表明,拟南芥中FT基因在叶片中表达,编码的FT蛋白可以从叶片运输到茎端,与FD(FLOWER INGLOCUSD)蛋白以复合体的形式激活花分生组织基因的表达而促进开花[25]。组成型表达TFL1能够延长植株的童期,使开花时间延迟[26]。通过RNAi技术将苹果植株内的TFL1表达量下调,苹果的童期明显缩短[27]。在梨中,TFL1基因作为花序分生组织特征基因,主要作用是延长植物的营养生长阶段,抑制成花转变[22]。本研究中,对照的TFL1基因的表达量在花芽分化时显著高于处理,而成花促进基因FT和LFY基因的表达量在花芽分化期均高于对照组,说明FT和LFY基因为成花促进基因,而TFL1基因为成花抑制基因,对这些成花基因表达的调控,将影响成花效果。本研究显示,对杂交后代的实生苗进行外源生长调节剂的喷施,将会促进成花转变,使实生苗提早开花,明显加速了育种进程,对果树育种有重要的意义。
| [1] | 陈东玫,李春敏,赵永波,等. 生长调节剂促进苹果实生树提早成花的研究[J]. 河北农业科学,2008,12(8):16-17[Chen D M,Li C M,Zhao Y B,et al. Research on the advanced flower of apple seedlings treated by growth regulators[J]. Journal of Hebei Agricutural Science,2008,12(8):16-17(in Chinese with English abstract)] |
| [2] | Chacko E K,Kohli R R,Swamy R D,et al. Growth regulators and flowering in juvenile mango(Mangifera indica L.)seedlings[J]. Symposium on Juvenility in Woody Perennials,1975,56:173-176 |
| [3] | 林晓东.激素调节花芽分化的研究进展[J].果树科学,1997,14(4):269-274[Lin X D. Advance in hormone′s regulation to flower bud differentiation[J]. Journal of Fruit Science,1997,14(4):269-274(in Chinese)] |
| [4] | 李学柱,邓 烈.喷布BA对柑桔内源激素及花芽分化的调控[J].园艺学报,1992,19(4):314-318[Li X Z,Deng L. Requlation of flower bud differentiation and the contents of endogenous hormones by spraying citrus trees with BA[J]. Acta Horticulturae Sinica,1992,19(4):314-318(in Chinese with English abstract)] |
| [5] | 过国南,郭兆年,阎淑芝.生长延缓剂和环剥对苹果杂种苗童期的影响[J].果树科学,1992,9(2):99-102[Guo G N,Guo Z N,Yan S Z. Effect of growth retardant and girdling to Juvenile phase of apple hybrid seedling[J]. Journal of Fruit Science,1992,9(2):99-102(in Chinese)] |
| [6] | 陈长宝,朱树华,周 杰.改良CTAB法提取成熟肥城桃果实的总RNA[J].山东农业科学,2009(5):102-104[Chen C B,Zhu S H,Zhou J. Improved CTAB method for total RNA extraction of mature Feicheng peach fruit[J]. Shandong Agricultural Sciences,2009(5):102-104(in Chinese with English abstract)] |
| [7] | Wu T,Zhang R,Gu C,et al. Evaluation of candidate reference genes for real time quantitative PCR normalization in pear fruit[J]. African Journal of Agricultural Research,2012,7:3701-3704 |
| [8] | 曹尚银,张俊昌,魏立华.苹果花芽孕育过程中内源激素的变化[J].果树科学,2000,17(4):244-248[Cao S Y,Zhang J C,Wei L H. Studies on changes of endogenous hormones in the differentiation period of flower bud in apple trees[J]. Journal of Fruit Science,2000,17(4):244-248(in Chinese with English abstract)] |
| [9] | 李晓旭,李家政.优化蒽酮比色法测定甜玉米中可溶性糖的含量[J].保险与加工,2013,13(4):24-27[Li X X,Li J Z. Determination of the content of soluble sugar in sweet corn with optimized anthrone colorimetric method[J]. Storage and Process,2013,13(4):24-27(in Chinese with English abstract)] |
| [10] | 鲁健章,孙丽华,周彦钢.凯氏定氮法测定鱼蛋白质含量的干扰因素分析[J].食品科学,2010,31(19):453-456[Lu J Z,Sun L H,Zhou Y G. Interference factors of protein determination in fish using Kjeldahl method[J]. Food Science,2010,31(19):453-456(in Chinese with English abstract)] |
| [11] | Bernier G,Havelange A,Houssa C,et al. Physiological signals that induce flowering[J]. Plant Cell,1993,5(10):1147-1155 |
| [12] | 李沛文,季作樑,梁立峰,等.荔枝大小年树营养芽及花芽分化与细胞分裂素的关系[J].华南农业大学学报,1985,6(3):1-8[Li P W,Ji Z L,Liang L F,et al. Relationship between vegetative and floral buds differentiation and CTK on Litchi alternate bearing trees[J]. Journal of South China Agricultural University,1985,6(3):1-8(in Chinese)] |
| [13] | 孙文全,褚孟嫄.梅树花芽生理分化期木质部液中赤霉素和细胞分裂素的变化[J]. 园艺学报,1988,15(2):73-76[Sun W Q,Chu M Y. The influence of cytokinin and gibberellin in the xylem SAP on floral initiation of Japanese apricot(Prunus mume Sieb.et Zucc)[J]. Acta Horticulturae Sinica,1988,15(2):73-76(in Chinese with English abstract)] |
| [14] | Grochowska M,Karaszewska A,Jankowska B,et al. The pattern of hormones of intact apple shoots and its changes after spraying with growth regulators[J]. Acta Horticulturae,1983,149:25-38 |
| [15] | 朴 宇,金德镐,曲柏宏.苹果梨幼树控旺促花方法研究[J].延边大学农学学报,1997,19(3):184-186[Piao Y,Jin D G,Qu B H. Study on methods of vigorous growing control and blossom acceleration for young‘Pingguo’ pear trees[J]. Journal of Yanbian University Agriculture Science,1997,19(3):184-186(in Chinese with English abstract)] |
| [16] | 卫晓铁,张明才,张 燕,等.乙烯利对不同基因型玉米节间伸长和內源激素的影响[J].农药学学报,2011,13(5):475-479[Wei X T,Zhang M C,Zhang Y,et al. Effects of ethephon on internode elongation and endogenous hormones in different genotypes of maize[J]. Chinese Journal of Pesticide Science,2011,13(5):475-479(in Chinese with English abstract)] |
| [17] | 黄羌维.龙眼内源激素变化和花芽分化及大小年结果的关系[J].热带亚热带植物学报,1996,4(2):58-62[Huang Q W. Changes in endogenous hormone contents in relation to flower bud differentiation and on-year or off-year fruiting of longan[J]. Journal of Tropical and Subtropical Botany,1996,4(2):58-62(in Chinese with English abstract)] |
| [18] | Kojima K. Changes of ABA,IAA and GAs levels in reproductive organs of citrus[J].JARQ,1997,31(4):271-280 |
| [19] | 李天红,黄卫东,孟昭清.苹果花芽孕育机理的探讨[J].植物生理学报,1996,22(3):251-257[Li T H,Huang W D,Meng Z Q. Study on the mechanisms of flower bud Induction in apple tree[J]. Acta Phytophsiologica Sinica,1996,22(3):251-257(in Chinese with English abstract)] |
| [20] | 郭金丽,张玉兰.苹果梨花芽分化期蛋白质、淀粉代谢的研究[J].内蒙古农牧学院学报,1999,20(2):80-83[Guo J L,Zhang Y L. A Study on the metabolism of protein and starch of apple-pear fruit tree in the stage of flower-bud differentiation[J]. Journal of Inner Mongolia Institute of Agriculture and Animal Husbandry,1999,20(2):80-33(in Chinese with English abstract)] |
| [21] | Pena L,Martín-Trillo M,Juárez J,et al. Constitutive expression of Arabidopsis LEAFY or APETALA 1 genes in citrus reduces their generation time[J]. Nature Biotechnology,2001,19(3):263-267 |
| [22] | 王 超,张全军,黄小三,等.‘金坠’梨成花相关基因PbLFY、PbTFL 1 和PbFT的克隆及时空表达模式分析[J].南京农业大学学报,2014,37(2):26-32. doi:10.7685/j.issn.1000-2030.2014.02.005[Wang C,Zhang Q J,Huang X S,et al. Cloning and temporal and spatial expression pattern of flowering-related genes PbLFY,PbTFL 1 and PbFT in‘Jinzhui’ pear[J].Journal of Nanjing Agricultural University,2014,37(2):26-32(in Chinese with English abstract)] |
| [23] | Endo T,Shimada T,Fujii H,et al. Ectopic expression of an FT homolog from citrus confers an early flowering phenotype on trifoliate orange(Poncirus trifoliata L.Raf.)[J].Transgenic Research,2005,14(5):703-712 |
| [24] | Hsu C Y,Liu Y,Luthe D S,et al. Poplar FT 2 shortens the juvenile phase and promotes seasonal flowering[J].Plant Cell,2006,18(8):1846-1861 |
| [25] | Corbesier L,Vincent C,Jang S,et al. FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis[J].Science,2007,316:1030-1033 |
| [26] | Alvarez-Buylla E,Benítez M,Espinosa-Soto C. Phenotypic evolution is restrained by complex developmental processes[J].HFSP Journal,2007,1(2):99-103 |
| [27] | Gyllenstrand N,Clapham D,Kllman T,et al. A Norway spruce FLOWERING LOCUS T homolog is implicated in control of growth rhythm in conifers[J].Plant Physiology,2007,144(1):248-257 |