
文章信息
- 盛超, 孟玲, 许庆辉, 李保平. 2015.
- SHENG Chao, MENG Ling, XU Qinghui, LI Baoping. 2015.
- 番茄和烟草对动植性烟盲蝽生长发育和捕食功能反应的影响
- Effects of tomato and tobacco plants on developmental performances and functional responses of zoophytophagous Nesidiocoris tenuis (Heteroptera:Miridae)
- 南京农业大学学报, 38(2): 261-266
- Journal of Nanjing Agricultural University, 38(2): 261-266.
- http://dx.doi.org/10.7685/j.issn.1000-2030.2015.02.013
-
文章历史
- 收稿日期:2014-07-20




植物是捕食性天敌昆虫捕食活动的小生境,它不仅影响猎物(植食性昆虫)的躲避和防卫行为[1],从而间接影响捕食性天敌的捕食效果;而且还可直接影响捕食性天敌的捕食行为和捕食效果[2]。此外,对于杂食性昆虫而言,植物又是其重要的营养来源之一[3],所以,植物对于杂食性昆虫,如盲蝽的捕食行为具有非常重要的影响[4]。
功能反应(functional response)是指每个捕食者的捕食率随环境中猎物密度变化的一种反应,即捕食者对猎物的捕食效应[5]。功能反应可用于估计由捕食而导致的猎物种群增长速率的降低趋势,可用于鉴别捕食者对猎物的选择性,以及描述群落内联结不同营养层级生物的食物网特征,也是生物防治中用于评价捕食性天敌控害作用的主要指标之一[6, 7]。研究捕食性昆虫的捕食功能反应试验常常在培养皿或类似的箱盒中进行[3],虽然这种方法便于比较不同研究对象的捕食作用,但研究结果与野外实际常常相差较大,尤其对于那些既取食植物又捕食其他昆虫的杂食性天敌昆虫难以揭示实际发生的捕食作用。
烟盲蝽(Nesidiocoris tenuis)是一种以肉食为主兼食植物的杂食性昆虫[8],由于烟盲蝽对微小体型害虫具有较好的控制作用,常作为优良天敌用于温室害虫的生物防治[9, 10, 11];研究[3]表明,杂食性盲蝽在作物大田中对害虫也具有较强的控制作用。迄今,烟盲蝽捕食桃蚜(Myzus persicae)、温室白粉虱(Trialeurodes aporariorum)、斜纹夜蛾(Spodoptera litura)低龄幼虫和小菜蛾(Plutella xylostella)幼虫等害虫的功能反应已有研究[12, 13, 14],且发现均符合Holling Ⅱ型模型[15]。但这些研究存在两点不足:一是均在封闭的小型容器(如培养皿)中进行试验;二是数据分析中所采用的Holling Ⅱ圆盘方程的假定与试验设计不符,该方程假定猎物密度水平始终一样[15],而试验在观察期(如:24 h)内均不能及时补充猎物使猎物密度始终保持一样。
因此,本研究首先采用非选择性饲喂试验,旨在明确斜纹夜蛾幼虫和植物(番茄和烟草)在烟盲蝽生长发育中的作用;然后,利用盆栽植物作为小生境,分别观察烟盲蝽在烟草和番茄上对不同密度斜纹夜蛾低龄幼虫的捕食效应,用Rogers[16]提出的Ⅱ型随机捕食者模型估计功能反应模型参数。 1 材料与方法 1.1 供试材料
供试烟草(Nicotiana tabacum)品种为‘中烟100’,于穴盘中育苗,当株高约5 cm时移入塑料杯(上口径7.5 cm、底径5 cm、高10 cm)中生长,待最大叶片长8 cm时供试。供试番茄(Solanum lycopersicum)品种为‘红光203’,于穴盘(长20 cm、宽30 cm)中播种育苗,幼苗长至高15 cm时供试。
供试斜纹夜蛾采自南京农业大学江浦农场大豆田,用人工饲料于室内继代饲养。供试烟盲蝽于2011年7月采自山东农业大学植物保护学院烟草试验田,采回后饲养在人工气候箱内的烟草植株上,饲喂斜纹夜蛾3龄幼虫(用前进行冰冻处理)作为动物性食物营养。用烟盲蝽雌虫进行试验。饲养条件为温度(26±1)℃、相对湿度(60±10)%、光/暗=14 h/10 h。 1.2 烟盲蝽在烟草和番茄上生长发育特征的观察
为揭示2种植物对烟盲蝽个体发育的影响,分别在有、无斜纹夜蛾幼虫的情况下将初孵若虫接到盆栽植物上,观察其生长发育至成虫羽化。将2~3对烟盲蝽成虫接到有斜纹夜蛾幼虫的烟草植株上,把烟草植株放于自制透明观察罩(长35 cm、宽20 cm、高35 cm)内1周,让成虫在烟草上产卵。1周后移除成虫,每日将烟草植株上刚孵化的单头烟盲蝽若虫用软毛笔移入不同处理的试管。管内先放入叶柄用棉球浸水保湿的待测植物叶片,再用镊子将斜纹夜蛾幼虫轻放在试管壁或植物叶片上,然后用纱布和橡皮筋封住试管口,放于人工气候箱内饲养。植物叶片隔天更换,斜纹夜蛾幼虫每日更换。每日观察若虫的发育与存活情况,记录羽化成虫的性别,将羽化成虫存放于低温冰箱内供测定和称干质量用。每处理重复25~30次。在体视显微镜(Motic 2.0成像软件)下测量成虫的后足胫节长度,并将10头为1组(共10组)的成虫放入烘箱内,在80 ℃下烘48 h后称干质量(Mettler AL204-1C电子天平);用后足胫节长度和体干质量分别代表成虫体型大小。 1.3 烟盲蝽在烟草和番茄上对斜纹夜蛾幼虫的捕食功能反应
将盆栽植物放入透明观察罩(长40 cm、宽25 cm、高30 cm)内,用软毛笔挑取斜纹夜蛾2龄幼虫置于植物叶片上,按照以下密度接虫:10、20、30、40、50和60头 · 株-1,猎物幼虫分布于2~3个叶片上,待幼虫开始取食(约2 h)后,取1头饥饿12 h的3日龄雌性烟盲蝽成虫放于其中1个叶片上;于12 h后移走烟盲蝽,计数剩余的猎物数量。每处理重复10次。 1.4 数据处理
用单因素方差分析比较在有、无斜纹夜蛾幼虫食物的情况下烟草与番茄之间的差异。比值数据经反正弦平方根转换后分析,以满足模型对分布型和方差齐次性的要求。 用logistic回归模型拟合被捕食猎物比例与猎物密度的关系,用基于最大似然估计的标准信息准则(AIC)在Holling Ⅱ与Ⅲ型模型之间进行选择,从而确定功能反应类型。由于本试验中在观察期内猎物密度随时间而降低,故采用Rogers[16]针对该类试验设计以Holling Ⅱ型功能反应模型为基础建立的Ⅱ型随机捕食者方程(type Ⅱ Rogers random predator equation):
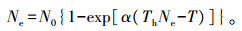
非选择性饲喂试验表明:在没有斜纹夜蛾幼虫食物的情况下,烟盲蝽靠取食植物叶片只能发育到3龄(番茄)或4龄(烟草);而在有斜纹夜蛾幼虫食物的情况下,烟盲蝽可以顺利发育到成虫(表 1)。在有斜纹夜蛾幼虫食物的处理下,烟盲蝽若虫发育历期和存活率在番茄与烟草之间没有显著差异(表 1);羽化率(图 1-A)和雌性比例(图 1-B)在番茄与烟草之间也没有显著差异;但子代雌成虫后足胫节长度(图 1-C)和体干质量(图 1-D)在两种植物处理间存在显著差异(P<0.05),取食烟草的子代雌成虫体型大于取食番茄的雌虫。
| d | ||||||
| 处理 Treatment |
1龄 1st instar |
2龄 2nd instar |
3龄 3rd instar |
4龄 4th instar |
5龄 5th instar |
若虫历期 Nymph time |
| 无幼虫食物Without larva food | ||||||
| 番茄Tomato | 3.54±0.22 | 4.11±0.56 | 2.67±1.20 | — | ||
| (34.62%) | (33.33%) | (0.00%) | ||||
| 烟草Tobacco | 3.96±0.19 | 3.50±0.48 | 4.00±0.41 | 1.50±0.50 | — | |
| (53.85%) | (28.57%) | (50.00%) | (0.00%) | |||
| 有幼虫食物With larva food | ||||||
| 番茄Tomato | 3.08±0.06 | 2.48±0.15 | 2.56±0.18 | 2.46±0.12 | 3.78±0.17 | 14.05±0.24 |
| (100.00%) | (96.00%) | (96.00%) | (96.00%) | (96.00%) | (84.00%) | |
| 烟草Tobacco | 3.40±0.12 | 2.90±0.15 | 2.14±0.11 | 2.40±0.17 | 2.95±0.19 | 13.53±0.32 |
| (100.00%) | (88.00%) | (96.00%) | (96.00%) | (100.00%) | (80.00%) | |
| 注:括号内数值为存活率。Values in brackets are survival rate. | ||||||
 | 图 1 烟盲蝽取食添加斜纹夜蛾幼虫的番茄和烟草叶片的发育表现 Fig. 1 Developmental performances of N.tenuis between feeding tobacco and tomato leaves as well as Spodoptera litura caterpillar*表示番茄与烟草处理间存在显著差异(P<0.05)。Columns with * above bars indicate significant differences between tomato and tobacco treatments. |
烟盲蝽分别在烟草和番茄植株上对不同密度斜纹夜蛾低龄幼虫的捕食反应均符合Holling Ⅱ功能反应模型的预测(适于Holling Ⅱ型功能反应的logistic回归;烟草:Z=13.03,P<0.001;番茄:Z=10.58,P<0.001)。在烟草上,当猎物密度从每株10头增大到每株30头时,被捕食的猎物数从平均(8.5±0.3)头增加到(20.0±0.9)头;当猎物密度继续增大到每株60头时,被捕食的猎物数仅多增加2~3头(图 2-A)。在番茄上,当猎物密度从每株10头增大到每株30头时,被捕食的猎物数从平均(7.9±0.2)头增大到(13.8±0.9)头;当猎物密度继续增大到每株60头时,被捕食的猎物数逐渐增加到(17.6±1.1)头(图 2-B)。在相同的猎物密度下在烟草上的被捕食猎物数量多于在番茄上的(图 2)。
 | 图 2 烟盲蝽在烟草(A)和番茄(B)植物上的被捕食数量与斜纹夜蛾低龄幼虫密度的关系 Fig. 2 Number of prey consumed by Nesidiocoris tenuis in relation to density of Spodoptera littura young larvae on plants of tobacco(A)and tomato(B)拟合线由Rogers Ⅱ型随机捕食者方程所得出,作图时为避免数据点重叠做了颤抖偏移。Lines are predicted by fitting the Rogers Ⅱ random predator equation. Jitter deviation was used to avoid overlapping data points. |
用非线性回归拟合Rogers Ⅱ型随机捕食者方程获得的参数估计值(表 2)表明:烟盲蝽在烟草上对斜纹夜蛾低龄幼虫的攻击率为3.056 h-1,处理猎物时间为0.034 h;在番茄上的攻击率为2.778 h-1,处理猎物时间为0.051 h;攻击率在烟草与番茄间没有显著差异(Z=1.51,P=0.13),猎物处理时间在烟草上显著短于在番茄上(Z=4.20,P<0.001)。
| 参数 Parameter |
估计值 Estimate |
标准误 SE |
置信域 95% CI |
Z值 Z-value |
P值 P-value |
| 烟草Tobacco | |||||
| 攻击率/h -1 Attack rate | 3.056 | 0.349 | 2.464~3.704 | 8.76 | <0.001 |
| 处理时间/h Handling time | 0.034* | 0.002 | 0.030~0.038 | 18.84 | <0.001 |
| 番茄Tomato | |||||
| 攻击率/h-1 Attack rate | 2.778 | 0.382 | 1.639~2.955 | 5.967 | <0.001 |
| 处理时间/h Handling time | 0.051 | 0.003 | 0.044~0.057 | 14.787 | <0.001 |
| 注:*表示参数在不同植物处理间存在显著差异(P<0.05)。* indicates significant differences between plants at 0.05 level. | |||||
本研究表明:斜纹夜蛾幼虫食物对于烟盲蝽发育至关重要;在有猎物幼虫的情况下,番茄和烟草叶片食物对烟盲蝽发育有一定的影响,取食烟草的子代成虫体型大于取食番茄的。说明烟盲蝽首先依赖于动物性营养,其次才是植物性营养。这与此前对烟盲蝽的研究结论是一致的[8, 11]。烟盲蝽不仅依赖于动物性营养,而且对取食的植物也存在选择性[21],其他杂食性盲蝽也有类似表现[22, 23]。这说明在利用烟盲蝽防治害虫的实践中,应注意作物种类的影响,在有利于烟盲蝽生长发育的作物上应用可望获得更好的防治效果。
本研究表明:植物影响烟盲蝽对斜纹夜蛾低龄幼虫的捕食功能反应,在烟草上的捕食效率高于在番茄上。这说明植物可影响烟盲蝽的捕食行为。关于植物小生境影响捕食行为的现象也存在于其他捕食性昆虫中[2]。
植物影响烟盲蝽捕食行为的原因可能有以下几方面:1)植物叶表物理特征可能影响烟盲蝽的搜寻和捕食行为。对某些瓢虫、草蛉、捕食性蝽和捕食螨的研究发现,植物表皮复杂的被毛或表皮蜡质可干扰昆虫的搜寻行为[24, 25, 26]。例如,Timms等[27]研究发现,2种瓢虫对在两种云杉叶上的蚜虫具有不同的功能反应表现。2)不同植物为烟盲蝽提供的营养差异可能影响烟盲蝽的捕食。烟盲蝽除主要捕食其他昆虫或螨外,还兼取食植物[8];若干研究发现烟盲蝽对不同植物存在取食选择性,而且取食不同植物对其产生的影响也不同[21, 23];类似这样的表现也存在于其他杂食性盲蝽中[4, 22]。3)斜纹夜蛾幼虫的取食可能诱导番茄产生对烟盲蝽取食的抗性。研究发现,植食性昆虫对植物的取食可诱导该植物产生次生防卫化学物质,从而影响杂食性盲蝽的取食[28]。本研究结果说明,在利用和评价烟盲蝽控害效果中,应予关注植物的影响。
山东农业大学植物保护学院郑方强、杨超和张小兵协助采虫,南京农业大学徐静参加部分试验,谨致谢意。
| [1] | Price P W, Bouton C E, Gross P, et al. Interactions among three trophic levels:influence of plants on interactions between insect herbivores and natural enemies[J]. Annual Review of Ecology and Systematics, 1980, 11:41-65 |
| [2] | Denno R F, Finke D, Langellotto G A. Direct and indirect effects of vegation structure and habitat complexity on predator-prey and predator-predator interactions[M]//Barbosa P, Catellanos I. Ecology of Predator-Prey Interactions. London:Oxford University Press, 2005:211-239 |
| [3] | Coll M, Guershon M. Omnivory in terrestrial arthropods:mixing plant and prey diets[J]. Annual Review of Entomology, 2002, 47:267-297 |
| [4] | Castañé C, Arnó J, Gabarra R, et al. Plant damage to vegetable crops by zoophytophagous mirid predators[J]. Biological Control, 2011, 59:22-29 |
| [5] | Murdoch W W, Briggs C J, Nisbet R M. Consumer-resource Dynamics[M]. Princeton:Princeton University Press, 2003:43 |
| [6] | Fernandez-Arhex V, Corley J C. The functional response of parasitoids and its implications for biological control[J]. Biocontrol Science and Technology, 2003, 13(4):403-413 |
| [7] | Fellowes M D E, van Alphen J J M, Jervis M A. Foraging behaviors[M]//Jervis M A. Insect as Natural Enemies, A Practical Perspective. London:Chapman and Hall, 2005:1-71 |
| [8] | Sanchez J A. Zoophytophagy in the plantbug Nesidiocoris tenuis[J]. Agricultural and Forest Entomology, 2008, 10:75-80 |
| [9] | 吴伟坚, 余金咏, 高泽正, 等. 杂食性盲蝽在生物防治上的应用[J]. 中国生物防治, 2004, 20(1):61-64 [Wu W J, Yu J Y, Gao Z Z, et al. The use of omnivorous mirids for biological control of pests[J]. Chinese Journal of Biological Control, 2004, 20(1):61-64(in Chinese with English abstract)] |
| [10] | Sanchez J A, Lacasa A, Arnó J, et al. Life history parameters for Nesidiocoris tenuis(Reuter)(Het., Miridae)under different temperature regimes[J]. Journal of Applied Entomology, 2009, 133:125-132 |
| [11] | Calvo J, Bolckmans K, Stansly P A, et al. Predation by Nesidiocoris tenuis on Bemisia tabaci and injury to tomato[J]. BioControl, 2009, 54:237-246 |
| [12] | 韦德卫, 贤小勇, 周至宏, 等. 烟盲蝽对斜纹夜蛾捕食功能反应的初步研究[J]. 河南农业大学学报, 1998, 32(1):55-59 [Wei D W, Xian X Y, Zhou Z H, et al. Preliminary study on the functional responses of Cyrtopeltis tenuis to Spodoptera litura[J]. Acta Agriculturae Universitatis Henanensis, 1998, 32(1):55-59(in Chinese with English abstract) |
| [13] | 官宝斌, 陈家骅, 陈乾锦, 等. 烟盲蝽对斜纹夜蛾幼虫和烟蚜的捕食功能反应[J]. 中国烟草学报, 1999, 5(4):21-24 [Gong B B, Chen J H, Chen Q J, et al. The predacions capacity and functional response of Cyrtopeltis tenuis reuter to aphidus and larvae of Prodenia liture(Fabiricius)[J]. Acta Tabacaria Sinica, 1999, 5(4):21-24(in Chinese with English abstract)] |
| [14] | 李令蕊, 薛仁风, 王晓婧, 等. 烟盲蝽对温室白粉虱和小菜蛾捕食作用研究[J]. 河北农业大学学报, 2008, 31(1):84-87 [Li L X, Xue R F, Wang X J, et al. Predation of Nesidiocoris tenuis to Trialeurodes vaporariorum and Plutella xylostella[J]. Journal of Agricultural University of Hebei, 2008, 31(1):84-87(in Chinese with English abstract)] |
| [15] | Holling C S. Some characteristics of simple types of predation and parasitism[J]. Canadian Entomologist, 1959, 91:385-398 |
| [16] | Rogers D. Random search and insect population models[J]. Jounal of Animal Ecology, 1972, 41:369-383 |
| [17] | Juliano S A. Nonlinear curve fitting:predation and functional response curves[M]//Scheiher S M, Gurevitch J. Design and Analysis of Ecological Experiments. London:Oxford University Press, 2001:178-196 |
| [18] | Bolker B M. Ecological Models and Data in R[M]. Princeton:Princeton University Press, 2008:268-270 |
| [19] | Bolker B M, Gardner B, Maunder M, et al. Strategies for fitting nonlinear ecological models in R, AD Model Builder and BUGS[J]. Methods in Ecology and Evolution, 2013, 4:501-512 |
| [20] | R Core Team. R:a language and environment for statistical computing[EB/OL]. Vienna, Austria:R Foundation for Statistical Computing, 2014. http://www.R-project.org/ |
| [21] | 路慧, 陈红印. 三营养级体系烟盲蝽对温室内不同作物种类的选择性[J]. 植物保护, 2007, 5(33):75-79 [Lu H, Chen H Y. Preference of omnivorous natural enemy Cyrtopeltis tenuis Reuter for plants in greenhouses in a tritrophic system[J]. Plant Protection, 2007, 5(33):75-79(in Chinese with English abstract)] |
| [22] | 吴伟坚, 余金咏, 梁广文. 盲蝽科昆虫的食性[J]. 昆虫知识, 2003, 40(2):108-111 [Wu W J, Yu J Y, Liang G W. Feeding habits of Miridae[J]. Entomological Knowledge, 2003, 40(2):108-111(in Chinese with English abstract)] |
| [23] | Ingegno B L, Pansa M G, Tavella L. Plant preference in the zoophytophagous generalist predator Macrolophus pygmaeus(Heteroptera:Miridae)[J]. Biological Control, 2011, 58:174-181 |
| [24] | Eigenbrode S D, Castagnola T, Rous M B, et al. Mobility of three generalist predators is greater on cabbage with glossy leaf wax than on cabbage with a wax bloom[J]. Entomologica Experimentalia et Applicata, 1996, 104:307-319 |
| [25] | Rutledge C E, Robinson A P, Eigenbrode S D. Effects of a simple plant morphological mutation on the arthropod community and the imapcts of predators on a principal insect herbivore[J]. Oecologia, 2003, 135:39-50 |
| [26] | Stavrinides M C, Skirvin D J. The effect of chrysanthemum leaf trichome density and prey spatial distribution on predation of Tetranychus urticae(Acari:Tetranychidae)by Phytoseileus persimilis(Acari:Phytoseiidae)[J]. Bulletin of Entomological Research, 2003, 93:343-350 |
| [27] | Timms J E, Oliver T H, Straw N A, et al. The effects of host plant on the coccinellid functional response:is the conifer specialist Aphidecta obliterata(L.)(Coleoptera:Coccinellidae)better adapted to spruce than the generalist Adalia bipunctata(L.)(Coleoptera:Coccinellidae)?[J]. Biological Control, 2008, 47:273-281 |
| [28] | Kaplan I, Thaler J S. Do plant defenses enhance or diminish prey suppression by omnivorous Heteroptera?[J]. Biological Control, 2011, 59:53-60 |