 2016, Vol. 52
2016, Vol. 52文章信息
- 蔡守平, 何学友, CeoffM Gurr, 曾丽琼, 黄金水, 嵇保中
- Shouping Cai, Xueyou He, M Gurr Ceoff, Liqiong Zeng, Jinshui Huang, Baozhong Ji
- 星天牛幼虫人工饲料的饲养效果及温度对其生长的影响
- Artificial Diet and Temperature Regimes for Successful Rearing of the Citrus Longhorned Beetle, Anoplophora chinensis (Coleoptera: Cerambycidae)
- ${metaVo.journalTitleCn}, 2016, 52(12): 141-149
- Scientia Silvae Sinicae, 2016, 52(12): 141-149.
- DOI: 10.11707/j.1001-7488.20161217
-
文章历史
- 收稿日期:2016-02-27
- 修回日期:2016-04-05
-
作者相关文章


2. 福建省林业科学研究院 国家林业局南方山地用材林培育重点实验室 福州 350012;
3. 福建农林大学应用生态研究所 福州 350002
2. Key Laboratory of Timber Forest Breeding and Cultivation for Mountainous Areas in Southern China of State Forestry Administration Fujian Academy of Forestry Fuzhou 350012 ;
3. Institute of Applied Ecology, Fujian Agriculture and Forestry University Fuzhou 350002
The citrus longhorned beetle, Anoplophora chinensis is widely distributed in China, Korea and Japan, with occasional records from Indonesia, Malaysia, Philippines, and Vietnam (Lingafelter et al., 2002; Hérard et al., 2006; Haack et al., 2010) [A. malasiaca was also placed in synonymy with A. chinensis (Lingafelter et al. 2002) , but A. malasiaca is still commonly used in Japan], however, breeding populations of A. chinensis had been found in France, Italy, and the Netherlands, as the insect spread mostly through international trade of live woody plants such as bonsai and nursery stock (Haack, 2006; Haack et al., 2010) . This cerambycid is a polyphagous pest that infests over 100 species of angiosperm tree and extends to conifers in the genera Cryptomeria (Cupressaceae) and Pinus (Pinaceae) . Larvae bore into the trunk through egg-laying scars and destroy the phloem and xylem. During its long period of development, this pest disrupts the vascular tissue resulting in structural weakness and can lead to tree death (Haack et al., 2010; Huang et al., 2012) . In China, A. chinensis is a particularly serious pest of citrus orchards (Gressitt, 1942; Xiao, 1992; Huang et al., 2012) and Casuarina equisetifolia L. forests (Huang et al., 1966; 2012) . In some areas, A. chinensis infestation rates can reach 50% resulting in afforestation failure (Huang et al., 1966; 2012) .
Developing control methods for this pest is a challenge because it is difficult to collect sufficient specimens for research, even from heavily infested orchards. As there is often only one larva at the base of each infected tree meaning that large numbers of trees need to be destructively sampled. Further, extracting the insect from woody tissue without injury is difficult and labour intensive. These practical factors add weight to the more general value of artificial diets in the study of arthropods in order to provide ready access to large numbers of insects of consistent age and physiological state. Several artificial diets and rearing protocols had been reported for Anoplophora species including A. glabripennis (Zhao et al., 1999; Dubois et al., 2002; Keena, 2005) and A. macularia (Thomson) (Lee et al., 1998) . However, no artificial diet has been reported for A. chinensis and the effects of temperature regime of development are not known. In this study, artificial diets and temperature regimes were compared with the objective of identifying optimal conditions for rearing A. chinensis.
1 Material and method 1.1 Diet preparationThe formulation of four artificial diets tested were showed in Tab. 1. These diets were mainly modified from diets developed by other researchers' studies on rearing cerambycid larvae (Lee et al., 1998; Zhao et al., 1999; Dubois et al., 2002) . Fresh C. equisetifolia tree logs without leaves were collected from Chihu Forest Farm (24°54′53″N, 118°54′14″E) , Hui'an County, Fujian Province, China. Short sections of log (less than 10 cm) , were dried at 55 ℃ for 12 h, and then sawn into sawdust using an electric saw. The sawdust was passed through a 3 mm sieve for use in selected diets. Agar and water were boiled and then mixed with the other ingredients to give four diets. Diets were dispensed into 40 mL plastic cups or 350 mL plastic jars to two-thirds capacity of the containers. Diet containers were autoclaved for 20 min at 121 ℃ and then stored at 4 ℃. Prior to use, the diets were warmed to room temperature and holes appropriate for the larva size were cut in the diet.
|
|
Adult A. chinensis were collected from a young C. equisetifolia plantation at the Chihu Forest Farm (see above) between late May and mid-June 2010. Adults were mated in a cage (40 cm×30 cm×60 cm) made of metal screening (0.8 mm in diameter) in the laboratory under unregulated conditions. Fresh twigs without leaves were provided as food. These stood in a glass beaker containing water and were replaced every two or three days. Bolts (40 cm long and ca. 5-8 cm in diameter) cut from C. equisetifolia trees were also provided as an oviposition substrate. These were put upright in a plastic beaker containing water and were replaced every three days (Fig. 1A) . The bark bearing eggs and larvae was removed from the bolts. Eggs were placed in Petri dishes (9 cm in diameter and 1.6 cm in depth) with moist filter paper in an unlit incubator (Fig. 1C) and larvae were used directly for rearing experiments. All experiments were carried out in the laboratory at Fujian Academy of Forestry, Fuzhou.

|
Fig.1 Laboratory rearing of A. chinensis on artificial diet A: Bolts of C. equisetifolia supplied for oviposition in metal cages; B: The bark of bolts with eggs or larvae was stripped; C: Eggs were put in Petri dish for hatching (arrows show larvae) ; D: Hole appropriate for the larval size was cut in the diet before rearing; E: Different rearing containers appropriate for the larval size were supplied; F: Pupal chamber made in the diet by larva; G: Pupae became dark gradually (commencing with eyes and mandibles) ; H: Whole pupa with uniform pigmentation, remaining in pupal chamber for 7-10 d; I: Adult crawled onto the surface of the diet. A: Bolts of C. equisetifolia supplied for oviposition in metal cages; B: The bark of bolts with eggs or larvae was stripped; C: Eggs were put in Petri dish for hatching (arrows show larvae) ; D: Hole appropriate for the larval size was cut in the diet before rearing; E: Different rearing containers appropriate for the larval size were supplied; F: Pupal chamber made in the diet by larva; G: Pupae became dark gradually (commencing with eyes and mandibles) ; H: Whole pupa with uniform pigmentation, remaining in pupal chamber for 7-10 d; I: Adult crawled onto the surface of the diet. |
Larvae were placed individually into 40 mL plastic cups of diets A-D with at least 20 replicates. Cups were placed in a fully randomized design within unlit incubator (meant the larvae were reared under dark condition) with unregulated, fluctuating temperature ranging from a monthly average low of approximately 10 ℃ in December-February (extreme low temperature in winter can reach as low as 0 ℃) and a maximum of 30 ℃ in June-August (extreme high temperature in summer can reach as high as 39 ℃) . Each larva was removed and placed into a new cup with fresh diet every two weeks. After two months, individual larvae were transferred to 350 mL plastic jars with diet. Diet was then replaced every month until pupation commenced in the cohort. After this time, larvae were checked every two days to determine pupation time. Pupae were left undisturbed in the containers to avoid damage from handling. Emergence date, any malformity and weight of each adult was recorded when they crawled onto the surface of the diet (without complementary nutrition) (Fig. 1I) , and then the adults from each diet type were pooled in metal cages to reproduce as described above. The second generation of larvae was reared on the same diet type as used for the earlier generation and, over the course of three years, the performance of three successive generations on each diet was assessed. Numbers of larvae used in each year are given in Tab. 2. About 30 larvae were reared in the first generation, except Diet A which had only 20 larvae because no enough larvae could be collected in a short time. For the second generation, 21 to 29 larvae were used for each diet. Because the emerged dates of adult was not synchronized, so only larvae were collected for rearing 7 to 10 days before the experiment. For the same reason, 12 to 20 larvae were reared in the third generation. Because insect performance was poor on Diet C, only two generations were reared. Larval numbers, duration time, pupation rate and adult emergence rate and weight were recorded. One-way analysis of variance (ANOVA) was used to test for effects of diet on larval and pupal periods. Two-way ANOVA was used to test for effects on adult weight between diets and generations. All multiple comparisons of means were performed using Tukey's test (SPSS 16.0) .
|
|
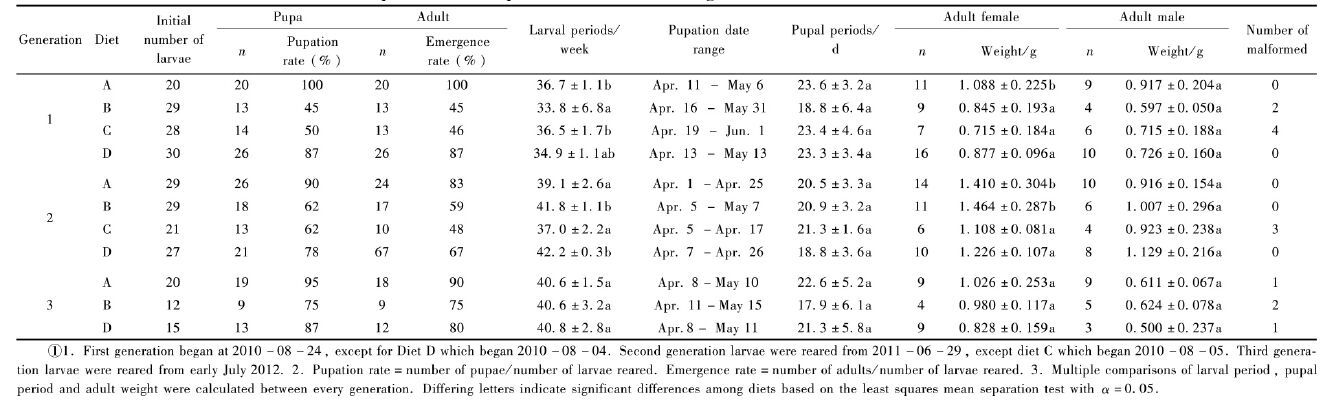
Development rates of A. chinensis on Diet A and D under different temperature regimes were studied in this experiment. Diet and insect preparation were as described in previous experiment whilst numbers of insects are given in Tab. 3. Containers with insects on Diet A and D were laid out in a fully randomized design within each of three unlit incubators (dark condition) with (i) fluctuating temperature, (ii) constant 25 ℃±1 ℃ and (iii) constant 28℃±1 ℃. Larvae were checked every week to determine survival rate. When pupation began, they were checked every two days to determine pupation time. Larval duration data for each diet and temperature regime were compared using ANOVA and multiple comparisons of means were performed using Tukey's test (SPSS 16.0) .
1.5 Weight and development of A. chinensis larvae on two superior diets under fluctuating and constant temperaturesThis experiment followed the protocol described in previous experiment, used Diets A and D and measured larval weight under fluctuating ambient temperature and constant 28 ℃. Fifty larvae (forty larvae for Diet D at 28℃) were assigned to each of five repeats on Diet A and D and laid out in a fully randomized design within an unlit incubator with fluctuating ambient temperature and an incubator at 28 ℃±1 ℃. Weight of every individual was recorded biweekly and the diet was replaced at the same time. After 12 weeks, larvae were transferred from 40 mL plastic cups to 350 mL plastic jars and larvae were weighed biweekly for 50 weeks or until pupation. For larvae that did not pupate within 50 weeks, larval weight was no longer recorded but the time of pupation and eventual adult emergence (or mortality) was recorded. Larval weigh data were subjected to repeated measures analysis of variance (ANOVA) using SPSS16.0 software. When a statistically significant difference between mean values was obtained, Tukey's test was applied (P=0.05) .
2 Result 2.1 Comparison of four diets over three successive A. chinensis generationsAll four diets led to at least some development to adult (Tab. 2) . Although some adults were malformed, the cohort from each diet treatment produced progeny that also completed a life cycle. There was significant different between different diets on pupation and emergence rates (one-way ANOVA, P<0.05) . Pupation and emergence rates on Diet A and D were significantly higher than other diets (Tab. 2) . Although the start date of rearing differed after the first generation according to the performance of cohorts on differing diets, pupation dates were approximately synchronized across treatments, all from early April to late May (Tab. 2) . When rearing time began in August (the first generation, as well as the second generation of Diet C) , the larvae took 34-37 weeks to pupate. When rearing began in late June or early July (Diet A, B, D for the second and third generation) , larval duration was about 40 weeks. Pupal duration for different diets and generations averaged between 18 to 23 days but individual insect pupal duration ranged widely from 10 to 34 days so there were no statistically significant differences between means (Tab. 2) . The mean weights of female adults were significantly different among diets and generations (two-way ANOVA, P<0.001 for generation and for diets) (Tab. 2) but there was no significant interaction (P>0.05) . The weights of male adults were different among generations (Tab. 2) with significant interaction between diet and generation (two-way ANOVA, P<0.001 for the generation and P=0.008 for the interaction, P>0.05 for the diets) . Means of female and male weight ranged from 0.715 g±0.184 g to 1.464 g±0.287 g (mean ± SE) , and 0.500 g±0.237 g to 1.129 g±0.216 g, respectively. Female adults on Diet A were heavier than those on other diets (except for the 2nd generation) , and weights of female adults on Diet C were less than those on other diets, but there was no obvious trend in the weights of male adults on different diets.
Before pupation, larvae constructed a pupal chamber in the diet (Fig. 1F) . After becoming uniformly dark in color, the pupa would remain in the pupal chamber for 7 to 10 days (Fig. 1H) after which the adult crawled to the surface of diet (Fig. 1I) .
2.2 Development of A. chinensis on two superior diets under fluctuating and constant temperaturesAll larvae fed and survived at first under all temperature regimes after they were transferred to the diet. At 24 weeks, larval survival was over 80% in all treatments except for Diet D under constant 28 ℃ on which survival was 72% (Tab. 3) . Under fluctuating temperatures, pupation was relatively synchronized, occurring between early April and middle May, similar to successive rearing experiment. Mean larval duration and pupation rate were 37.7 weeks ±1.2 weeks and 90% for Diet A, 39.6 weeks±2.2 weeks and 83% for Diet D. Under constant temperatures, larval duration was more variable and significantly longer (one-way ANOVA, P<0.05) than under fluctuating temperatures and mortality was relatively high (Tab. 3) . The rate of adult malformation was higher under constant rather than fluctuating temperature. At constant 25 ℃, larvae took an average of 41.5 weeks±4.6 weeks and 40.3 weeks±3.4 weeks to become pupae on Diet A and D, respectively. At constant 28 ℃, the corresponding means were 51.4 weeks±6.1 weeks and 56.9 weeks±7.4 weeks. Pupation rates were lowest (50% and 20% on Diet A and D respectively) under constant 28 ℃. The incidence of adult malformation under constant was higher than fluctuating temperature.
|
|
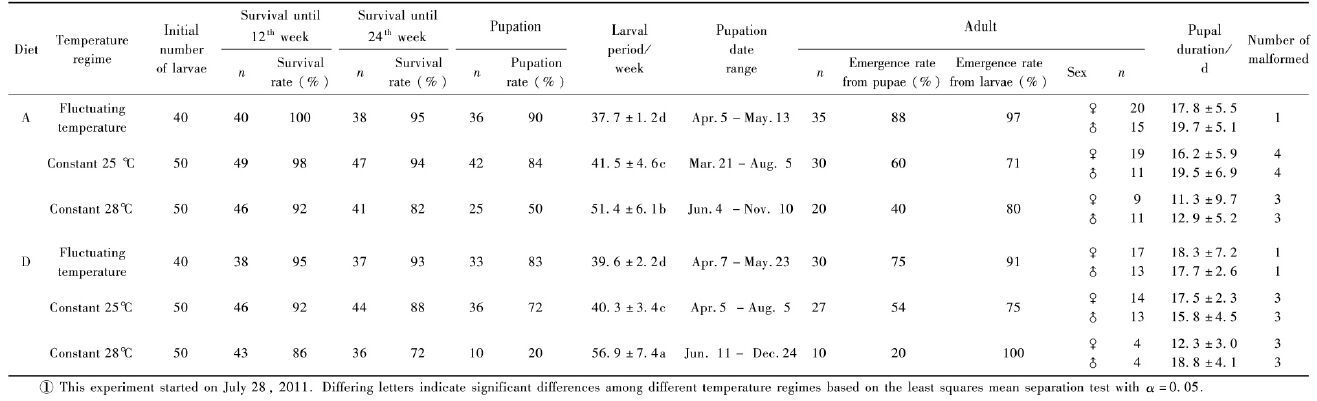
Every larva was taken out for recording weight every two weeks, and then put back to the diet. There was no difference in initial larval weight among diets and temperature regime treatments (one-way ANOVA, P>0.05) (Fig. 2) . Thereafter, larval weights on diet D increased more rapidly than on Diet A under both temperature regimes. Under fluctuating temperatures, larval weights peaked at 16 weeks (in late November) , at (2.077±0.185) g on Diet D, which was significantly heavier than that on Diet A at (1.685±0.131) g (P<0.05) . After 16 weeks, larval weight began to decline until pupation or death for both Diet A and D. Under fluctuating temperatures, pupation on Diet A began after 36 weeks, with an average larval duration of (37.9±1.5) weeks. Pupation was relatively synchronized, commencing in week 36, with all larvae pupated or dead by week 41 and a pupation rate of 84% (Tab. 4) . On Diet D, however, no larvae pupated and larval weight sustained a downward trend until death. Under constant 28 ℃, larval weight peaked at 2.106 g±0.183 g at week 16 on Diet D and 1.785 g±0.129 g at week 20 on Diet A, respectively, and there was a significant difference between the maximal weights (P<0.05) . After that, larval weight declined until pupation or death (Fig. 2) . The pupation rate was 56% on Diet A, but insects took longer to pupate, larval duration ranging widely from 36 to 55 weeks with an average of 46.7 weeks±5.8 weeks. On Diet D, only one larva pupated, at week 40.6, giving a pupation rate of 2.5% (Tab. 4) . It seemed that larvae on Diet D were more vulnerable to disturbance (larvae were periodically taken out for measuring weight) .

|
Fig.2 Weight dynamic of A. chinensis larvae on diets A and D under fluctuating and constant temperature |
|
|
Three consecutive generations of A. chinensis were successively reared on artificial diets. The majority of adults reared from artificial diets exhibited apparently normal behavior and morphology, as is reflected in the multiple generations produced. There were, however, marked differences in performance between diet types and rearing conditions. Performance of insects was especially good on Diets A and D that contained C. equisetifolia sawdust and wheat bran (and other ingredients) . Omission of either of these major ingredients led to significantly poorer insect performance. The influence of host plant tissue in artificial diets on the development of other cerambycids varies markedly, being of no benefit to detrimental in some cases. Harley et al. (1968) found that the addition of host plant tissue to the diet did not increase the development rate of Cerambycidae larvae. Payne et al. (1975) found that the early growth rate of Prionus imbricornis larvae was greatest when alphacel was substituted for the wood of pecan. Lee et al. (1998) found that addition of citrus (host plant) sawdust did not increase the survival rate of A. macularia larvae. Other studies, however, demonstrated that the addition of host plant tissue to artificial diets may be important for larval development. For example, when rearing Japanese sawyer, Monochamus alternatus Hope, the addition of inner bark in yeast-containing artificial diets led to increased pupation rate and pupal weight (Kosaka et al., 1990) .Aloo et al. (1994) also reported that inclusion of host plant tissue had a significant influence on larval growth and development of M. alternatus. Gindin et al. (2009) reported that larvae of Capnodis tenebrionis fed on any diet lacking cortex tissue of plum or peach trees survived for a relatively short period (maximum 5-6 weeks) and never reached the pupal stage. Gardiner (1970) found that the addition of pulverized plant material increased the acceptance of the diet by young Cerambycidae larvae. In the present study, the combined addition of C. equisetifolia sawdust and wheat bran significantly increased pupation and emergence rates compared to Diet B and C that lacked either C. equisetifolia sawdust or wheat bran. Sawdust of C. equisetifolia in the diets may act as a phagostimulant by virtue of its physical texture in addition to its chemical composition and nutritional value. The role of wheat bran is unclear but it too is likely to have significance as a nutrient source and possibly significant in providing physical stimulation.
Irrespective of diet, temperature had a strong effect on the development of A. chinensis. Temperature is believed to be important factor regulating Anoplophora spp. larval development (Adachi, 1994; Zhao et al., 1999; Keena et al., 2010) . At 20 ℃, most larvae of A. glabripennis pupated without either a chill period or exposure to higher temperatures (Keena et al., 2010) . In contrast, few larvae pupated at 25 ℃, and no larvae at 30 ℃, in the absence of a chill period, and the timing of the chill period determined the instar at pupation. Whilst that study illustrated the importance of low temperature in that species, a more general requirement for pupation in this genus is suggested by work with A. glabripennis. Zhao et al. (1999) demonstrated that a low temperature (5 ℃) for three months decreased the larval duration, increased pupation rate, and synchronized development time, although a larval chill period during development was not considered essential by those authors. Another conspecific, A. malasiaca, failed to pupate at a constant 25 or 30℃, whilst under fluctuating temperatures, more than 70% of larvae survived and completed their life cycle in 1 or 2 years (from eggs to adult emergency) (Adachi, 1994) .
In the present study, the pupation and emergence rate under fluctuating temperatures was higher than that under a constant temperature on either of the superior diets. The pupation time of larvae under fluctuating temperatures was more tightly synchronized than under constant temperature, between early April and late May, which is consistent with pupation under field conditions (Xiao, 1992; Huang et al., 2012) . Under constant temperatures (25 and 28 ℃) , pupation timing was extended and highly variable. A higher adult malformation rate was also observed. Results show that temperature conditions affect the determination of the timing of A. chinensis pupation and adult emergence and this is consistent with other studies of congeneric. Although a larval chill period is not essential (a proportion of larvae pupated under constant temperature) , it does optimize rearing. Performance in the present study under fluctuating temperatures, when the average monthly temperature in our laboratory is approximately 10 ℃ in December, January and February and can fall to 0 ℃ (the larvae get their peak weights before this time) , increased the pupation rate and led to adult emergence at the normal time of the year. However, the effects of different chill regimes on development of A. chinensis merit further study in order to optimize the performance of cultures. And the time of chill period and its coevolution relation with the larvae weight need more deeply evaluated. Meanwhile, the reason why the pupation rates were very low on Diet D whether under fluctuating or constant 28 ℃ after the larvae were periodically taken out for weighing was not very clear. In conclusion, the present body of results provides a valuable resource for culturing A. chinensis, specifically by identifying suitable artificial diets and the temperature effects on larval development.
| [] | Adachi I.1994. Development and life cycle of Anoplophora malasiaca (Thomson) (Coleoptera:Cerambycidae) on citrus trees under fluctuating and constant temperature regimes. Appl Entomol Zool , 29 (4) : 485–497. |
| [] | Aloo I, Katagiri K.1994. Assessment of artificial rearing method for the pine sawyer beetle, Monochamus alternatus (Coleoptera:Cerambycidae). Bull. Tokyo Univ For , 92 : 153–166. |
| [] | Dubois T, Hajek A E, Susan S.2002. Methods for rearing the Asian longhorned beetle (Coleoptera:Cerambycidae) on artificial diet. Ann Entomol Soc Am , 95 (2) : 223–230. DOI:10.1603/0013-8746(2002)095[0223:MFRTAL]2.0.CO;2 |
| [] | Gardiner L M.1970. Rearing wood-boring beetles (Cerambycidae) on artificial diet. Can Entomol , 102 (1) : 113–117. DOI:10.4039/Ent102113-1 |
| [] | Gindin G, Kuznetsova T, Protasov A, et al.2009. Artificial diet for two flat-headed borers, Capnodis spp. (Coleoptera:Buprestidae). Eur J Entomol , 106 (4) : 573–580. DOI:10.14411/eje.2009.072 |
| [] | Gressitt J L.1942. Destructive long-horned beetle borers at Canton, China. Lingnan Nat Hist Surv Mus Spec Pub : 1–60. |
| [] | Haack R A.2006. Exotic bark and wood-boring Coleoptera in the United States:recent establishments and interceptions. Can J For Res , 36 (2) : 269–88. DOI:10.1139/x05-249 |
| [] | Haack R A, Hérard F, Sun J, et al.2010. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle:a worldwide perspective. Annu Rev Entomol , 55 : 521–546. DOI:10.1146/annurev-ento-112408-085427 |
| [] | Harley K L S, Willson B W.1968. Propagation of a cerambycid borer on a meridic diet. Can J Zool , 46 (6) : 1265–1266. DOI:10.1139/z68-174 |
| [] | Hérard F, Ciampitti M, Maspero M, et al.2006. Anoplophora species in Europe:infestations and management processes. EPPO Bull , 36 (3) : 470–474. DOI:10.1111/epp.2006.36.issue-3 |
| [] | 黄 炳勋, 陈 清机, 吕 飞平.1966. 木麻黄害虫星天牛(Anoplophora chinensis (Forster))的习性及防治经验. 林业科学 , 11 (2) : 146–147. |
| [] | 黄 金水, 何 学友. 2012. 中国木麻黄病虫害. 北京: 中国林业出版社 : 79 -107. |
| [] | Keena M A.2005. Pourable artificial diet for rearing Anoplophora glabripennis (Coleoptera:Cerambycidae) and methods to optimize larval survival and synchronize development. Ann Entomol Soc Am , 98 (4) : 536–547. DOI:10.1603/0013-8746(2005)098[0536:PADFRA]2.0.CO;2 |
| [] | Keena M A, Moore P M.2010. Effects of temperature on Anoplophora glabripennis (Coleoptera:Cerambycidae) larvae and pupae. Environ Entomol , 39 (4) : 1323–1335. DOI:10.1603/EN09369 |
| [] | Kosaka H, Ogura N.1990. Rearing of the Japanese pine sawyer, Monochamus alternatus (Coleoptea:Cerambycidae), on artificial diets. Appl Entomol Zool , 25 (4) : 532–535. |
| [] | Lee C Y, Lo K C.1998. Rearing of Anoplophora macularia (Thomson) (Coleoptera:Cerambycidae) on artificial diets. Appl Entomol Zool , 33 (1) : 105–109. |
| [] | Lingafelter S W, Hoebeke E R. 2002. Revision of Anoplophora (Coleoptera:Cerambycidae). Washington, DC:Entomological Society of Washington, 236. |
| [] | Payne J A, Lowman H, Pate R R.1975. Artificial diets for rearing the tilehorned Prionus. Ann Entomol Soc Am , 68 (4) : 680–682. DOI:10.1093/aesa/68.4.680 |
| [] | 萧 刚柔. 1992. 中国森林昆虫. 北京: 中国林业出版社 : 444 -445. |
| [] | 赵 军, 小仓 信夫, 矶野 昌弘.1999. 光肩星天牛的人工饲养(Ⅰ). 北京林业大学学报 , 21 (4) : 58–61. |