 2012, Vol. 48
2012, Vol. 48文章信息
- 江海澜, 王俊刚, 邓小霞, 彭俊, 马天文, 何泽敏
- Jiang Hailan, Wang Jungang, Deng Xiaoxia, Peng Jun, Ma Tianwen, He Zemin
- 梭梭漠尺蛾幼虫取食后梭梭的生理响应
- Physiological Responses of Haloxylon ammodendron to feeding of Desertobia heloxylonia
- 林业科学, 2012, 48(10): 170-173.
- Scientia Silvae Sinicae, 2012, 48(10): 170-173.
-
文章历史
- 收稿日期:2011-09-18
- 修回日期:2012-04-06
-
作者相关文章



梭梭(Haloxylon ammodendron)为灌木或小乔木,广泛分布于亚、非荒漠区(蒋进, 1992),分布在我国35°—48°N、60°—111°E之间的干旱沙漠地带,种类主要为梭梭和白梭梭(H. persicum)(张景波等, 2010)。我国现有梭梭林约1 156.72万hm2,其中新疆占72.3%,大多分布在准噶尔盆地古尔班通古特沙漠。梭梭具有较强的耐旱性和耐盐性(周智彬等,2002;赵明范等,1997),对荒漠区防风固沙、减小风速、改造沙区气候、保障分布区农业生产和生态平衡有重要意义(张霞等,2006;李霞等,2009a; 2009b)。
梭梭漠尺蛾(Desertobia heloxylonia)为近年发生于新疆北疆的突发性新害虫(薛大勇等,2006),1年发生1代,以蛹越夏越冬;日平均气温达10℃左右时,越冬代蛹开始羽化;其幼虫取食梭梭、白梭梭、白皮沙拐枣(Caligonum leucocladum)及沙拐枣(C. mongolicum)等旱生植物(刘茂秀等,2005),在准噶尔盆地南缘的古尔班通古特沙漠主要取食梭梭(李亦松等,2007)。
害虫与寄主植物的关系是昆虫生态学与植物生理学的研究重点。一般认为,昆虫的取食对植物生长发育不利,植物会被动地产生各种次生性物质以协调生长或进行防御(王丽艳等,2011;陈建明等,2000),也能凭借其健壮的生长和繁殖机能来忍受或补偿所受到的伤害(孟凡立等,2011;Simms,2000)。梭梭在逆境条件下的适应性研究报道甚多(谭德远等,2004a;2004b;郭泉水等,2004;2005),但尚未涉及害虫胁迫下梭梭的生理响应。鉴于此,本文研究了梭梭漠尺蛾幼虫取食梭梭后其植株体内叶绿素、可溶性糖、可溶性蛋白、游离脯氨酸和丙二醛含量的变化,以期揭示梭梭漠尺蛾幼虫取食胁迫下梭梭所发生的生理响应。
1 材料与方法 1.1 试验材料1) 供试虫源 试验用梭梭漠尺蛾幼虫采自新疆准噶尔盆地南缘的古尔班通古特沙漠边缘。于人工智能气候箱中(温度为28℃,湿度为70%)用新鲜梭梭同化枝条饲养,待幼虫发育至3~4龄时用于试验。
2) 供试梭梭 石河子大学农学院试验站(86°E, 44°N)沙漠沙土栽培的4年生梭梭。
1.2 试验方法1) 试验设置方法 取长势一致的梭梭植株,分别在其同一部位的同化枝上接0,1,3,5,7,9头梭梭漠尺蛾幼虫并用细纱网罩住,然后用细纱网将整株梭梭罩住;于24 h后在同一株接虫和未接虫的同化枝上分别取样。各处理重复3次;样品用预冷的冰盒带回实验室测定其生理指标。
2) 生理指标测定方法 叶绿体色素含量采用乙醇-丙酮混合浸提法提取,测定方法参照李玲(2009);可溶性糖含量测定采用蒽酮比色法;可溶性蛋白质含量测定采用考玛斯亮蓝(G-250比色法)(王学奎,2006);游离脯氨酸含量测定采用酸性茚三酮法;丙二醛(MDA)含量测定采用硫代巴比妥酸法(郑炳松,2006)。
1.3 数据统计与分析使用Excel进行数据处理,采用SPSS17.0统计软件进行Duncan法方差分析和多重比较。
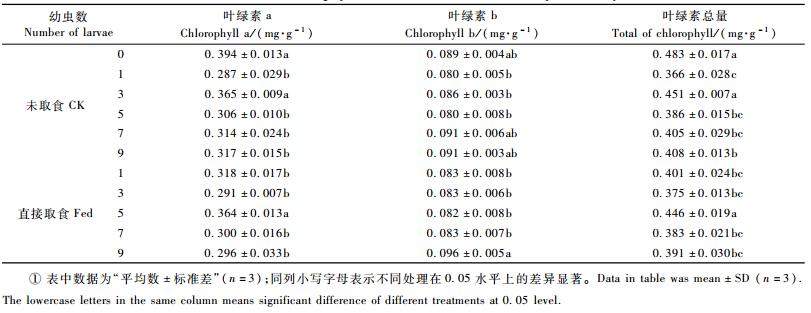
2 结果与分析 2.1 梭梭漠尺蛾幼虫取食对梭梭叶绿素含量的影响对各处理梭梭的叶绿素a、b及叶绿素总含量进行了测定,结果见表 1。梭梭漠尺蛾幼虫取食24 h后,梭梭叶绿素a、b及叶绿素总量均出现下降;叶绿素a和叶绿素总量除了未取食(接虫3头)处理和直接取食(接虫5头)处理外,其他处理与对照差异显著;而叶绿素b含量除了直接取食(接虫9头)处理外,其他处理之间差异不显著;说明梭梭漠尺蛾幼虫的取食对梭梭的叶绿素含量的影响不严重。
|
|
如图 1所示:梭梭漠尺蛾幼虫取食胁迫24 h后,梭梭可溶性糖含量的变化差异显著。除了处理7未直接受害的同化枝可溶性糖含量与对照差异不显著外,其他处理均显著高于对照;处理3梭梭受害同化枝的可溶性糖含量显著高于其他处理且比对照增加了85.74%,而未直接受害同化枝条的可溶性糖含量却只比对照增加了18.24%。但梭梭枝条内可溶性糖含量与为害梭梭漠尺蛾幼虫种群数量无相关性,这是否表明植株个体间存在抗性差异,尚待研究。

|
图 1 梭梭漠尺蛾幼虫取食后梭梭可溶性糖含量的变化 Fig.1 Content of soluble sugar in H. ammodendron induced by D. heloxylonia |
如图 2所示:梭梭漠尺蛾幼虫取食胁迫24 h后,梭梭可溶性蛋白含量的变化差异显著。接虫植株的可溶性蛋白含量随接虫数量的增多先降低后增多;接种1头幼虫同化枝的可溶性蛋白含量与对照差异不显著,与对照比较只下降了10.83%,接种9头幼虫的被食和未受害同化枝的可溶性蛋白含量分别增加了153.31%和135.86%。

|
图 2 梭梭漠尺蛾幼虫取食后梭梭可溶性蛋白含量的变化 Fig.2 Content of soluble protein in H. ammodendron induced by D. heloxylonia |
如图 3所示:梭梭漠尺蛾幼虫取食胁迫24 h后,游离脯氨酸含量的变化差异显著,接种9头及5头和7头的未直接受害梭梭枝叶的游离脯氨酸含量与对照差异显著,其他接种幼虫的梭梭同化枝游离脯氨酸含量均与对照差异不显著。其中,接种1头幼虫的未直接受害同化枝的游离脯氨酸含量只比对照降低了13.48%,接种7头幼虫的未直接受害同化枝则比对照上升了95.65%。

|
图 3 梭梭漠尺蛾幼虫取食后梭梭游离脯氨酸含量的变化 Fig.3 Content of free praline in H. ammodendron induced by D. heloxylonia |
如图 4所示:梭梭漠尺蛾幼虫取食胁迫24 h后,丙二醛含量的变化差异显著。梭梭同化枝的丙二醛含量随着接种幼虫数量的增加而增加,各处理的梭梭同化枝丙二醛含量均显著高于对照。其中,以接种9头幼虫未直接受害的梭梭同化枝的丙二醛含量最高,比对照增加了103.97%,而接种5条的直接受害同化枝的丙二醛含量增加了81.07%。

|
图 4 梭梭漠尺蛾幼虫取食后梭梭丙二醛含量的变化 Fig.4 Content of MDA in H. ammodendron induced by D. heloxylonia |
植物受昆虫取食后在个体发育、营养成分、次生物质代谢等方面会发生防御变化,并且这种诱导抗虫性能在植物体内表现出系统性的传递。在逆境条件下植物主要通过主动积累有机和无机物如可溶性糖、可溶性蛋白质以及游离氨脯氨酸等,以提高自身细胞液的浓度,降低渗透势、维持细胞正常的膨压等生理过程(孙存华等,2007)。植物在被害虫取食、干旱以及低温等情况下常主动积累一些可溶性糖;游离脯氨酸则能保持膜结构的完整性、保护细胞和酶的空间结构、参与解毒及增强膜中蛋白质分子间的水合力(周运刚等,2010),可溶性蛋白质可增强细胞的抵抗能力(郭丽等, 2010)。因此,植物受逆境胁迫的程度可以通过测定植物的渗透调节物质含量变化进行验证。
试验发现梭梭漠尺蛾幼虫取食处理能诱导梭梭植株直接被食和未直接被食部位的可溶性蛋白、可溶性糖含量增加、未直接被食部位的游离脯氨酸含量增高,尤其以接种9头幼虫时梭梭植株渗透调节物质积累最多。已有研究表明作物受害虫为害后抗虫品种体内可溶性总糖含量会明显增加(李进步等, 2008)。这表明梭梭漠尺蛾幼虫取食所诱导的梭梭体内这3种物质的积累或增加是其诱导抗虫性的系统诱导反应。但试验也同时发现:在接种1头梭梭漠尺蛾幼虫时梭梭体内可溶性蛋白含量和游离脯氨酸含量出现下降且与对照差异不显著,这表明梭梭对少量梭梭漠尺蛾幼虫的取食危害具有一定的容忍能力, 这种能力的获得可能是梭梭与梭梭漠尺蛾长期协同进化的结果。
植物依靠叶绿素进行光合作用,叶绿素的破坏或降解都会直接导致植物光合速率的降低,而植物光合作用进程或光合作用强度都会受到外界条件的影响(陈建明等,2003;苏培玺等,2006)。试验发现梭梭漠尺蛾幼虫取食危害24 h后梭梭植株的叶绿素a、b和叶绿素总量的含量有不显著的下降趋势, 这表明受害的梭梭叶片光合作用能力也有所变化。
正常状态下植物体内活性氧的产生和清除处于相对动态平衡中,但在逆境条件下其体内活性氧自由基产生的速度超出了植物自身清除的能力,进而发生膜脂过氧化作用产生丙二醛;植物受逆境胁迫时的MDA含量是植物膜系统稳定与否或反映膜损伤程度的重要指标(江海澜等,2011;潘晓云等,2002)。在受到梭梭漠尺蛾幼虫取食危害逐步加重时,梭梭植株MDA含量呈现上升趋势。接种1头幼虫时梭梭的MDA含量与对照差异不显著,表明这时的取食胁迫并不没有超出梭梭体内代谢所容忍的范围;而在幼虫数量增加时MDA含量显著上升,说明过多的取食危害会诱导梭梭体内产生多余的活性氧自由基,但其是否对梭梭的膜系统造成了损害还有待研究。
| [] | 陈建明, 俞晓平, 葛秀春, 等. 2000. 水稻植株防御白背飞虱为害的某些生理反应. 中国水稻科学, 14(1): 43–47. |
| [] | 陈建明, 俞晓平, 陈俊伟, 等. 2003. 水稻植株光合作用能力的变化与其抗白背飞虱的关系. 核农学报, 17(6): 423–426. |
| [] | 郭丽, 贾秀领, 张凤路, 等. 2010. 定位水氮组合对冀5265小麦叶片硝酸还原酶、可溶性蛋白及产量的影响. 华北农学报, 25(1): 180–184. DOI:10.7668/hbnxb.2010.01.036 |
| [] | 郭泉水, 谭德远, 刘玉军, 等. 2004. 梭梭对干旱的适应及抗旱机理研究进展. 林业科学研究, 17(6): 796–803. |
| [] | 郭泉水, 谭德远, 王春玲, 等. 2005. 接种肉苁蓉对梭梭天然林的影响研究. 生态学杂志, 24(8): 867–871. |
| [] | 江海澜, 王俊刚, 邓小霞, 等. 2011. 草甘膦对龙葵苗期生理指标的影响. 西北农业学报, 20(6): 186–189. |
| [] | 蒋进. 1992. 极旱环境中两种梭梭蒸腾的生理生态学特点. 干旱区研究, 9(4): 14–17. |
| [] | 李进步, 方丽平, 吕昭智, 等. 2008. 棉花抗蚜性与可溶性糖含量的关系. 植物保护, 34(2): 26–30. |
| [] | 李玲. 2009. 植物生理学模块实验指导. 北京, 科学出版社.. |
| [] | 李霞, 马永清, 宋玉霞, 等. 2009a. 肉苁蓉寄生对梭梭幼苗水分生理特征及生长的影响. 西北林学院学报, 24(3): 20–23. |
| [] | 李霞, 马永清, 宋玉霞, 等. 2009b. 肉苁蓉寄生对梭梭幼苗保护酶活性及渗透调节物质的影响. 生态学杂志, 28(8): 1531–1536. |
| [] | 李亦松, 张建华, 李小燕, 等. 2007. 新疆准噶尔盆地南缘梭梭漠尺蛾生物学特性的初步研究. 新疆农业科学, 44(6): 779–781. |
| [] | 刘茂秀, 丛者福, 魏岩. 2005. 莫索湾地区梭梭群落种间关联性的研究. 石河子大学学报:自然科学版(4): 449–453. |
| [] | 孟凡立, 王志坤, 孙晶, 等. 2011. 蚜虫取食大豆诱导大豆异黄酮变化的规律. 作物杂志, 1: 59–62. DOI:10.3969/j.issn.1001-7283.2011.01.016 |
| [] | 潘晓云, 曹琴东, 王根轩. 2002. 膜脂过氧化作为扁桃品种抗寒性鉴定指标研究. 生态学报, 22(11): 1902–1912. DOI:10.3321/j.issn:1000-0933.2002.11.015 |
| [] | 苏培玺, 严巧娣. 2006. C4荒漠植物梭梭和沙拐枣在不同水分条件下的光合作用特征. 生态学报, 26(1): 75–82. |
| [] | 孙存华, 李扬, 贺鸿雁, 等. 2007. PEG6000渗透胁迫对藜幼苗叶片渗透调节物质的影响. 安徽农业科学, 35(25): 7784–7786. DOI:10.3969/j.issn.0517-6611.2007.25.027 |
| [] | 谭德远, 郭泉水, 王春玲. 2004a. 我国肉苁蓉资源状况及开发利用研究. 林业资源管理, 2: 29–32. |
| [] | 谭德远, 郭泉水, 王春玲, 等. 2004b. 寄生植物肉苁蓉对寄主梭梭生长及生物量的影响研究. 林业科学研究, 17(4): 472–478. |
| [] | 薛大勇, 施登明, 邢海洪, 等. 2006. 漠尺蛾属研究及一新种记述(鳞翅目, 尺蛾科, 灰尺蛾亚科). 动物分类学报(1): 193–199. |
| [] | 王丽艳, 张海燕, 朱莹, 等. 2011. 水稻对稻负泥虫取食的生理响应. 应用昆虫学报, 48(4): 928–933. DOI:10.7679/j.issn.2095-1353.2011.155 |
| [] | 王学奎. 2006. 植物生理生化实验原理和技术. 北京, 高等教育出版社. |
| [] | 张景波, 王葆芳, 郝玉光, 等. 2010. 我国梭梭林地理分布和适应环境及种源变异. 干旱区资源与环境, 24(5): 166–171. |
| [] | 张霞, 邓必建, 姚新花, 等. 2006. 不同温度条件下PEG引发梭梭种子对其幼苗生理生化的影响. 种子, 25(12): 5–7. DOI:10.3969/j.issn.1001-4705.2006.12.002 |
| [] | 赵明范, 葛成, 翟志中. 1997. 干旱地区次生盐碱地主要造林树种抗盐指标的确定及耐盐能力排序. 林业科学研究, 10(2): 194–198. |
| [] | 郑炳松. 2006. 现代植物生理生化研究技术. 北京, 气象出版社. |
| [] | 周运刚, 王俊刚, 马天文, 等. 2010. 不同DPC (缩节胺)处理对棉花生理生化特性的影响. 新疆农业科学, 47(6): 1142–1146. DOI:10.6048/j.issn.1001-4330.2010.06.017 |
| [] | 周智彬, 徐新文. 2002. 塔克拉玛干沙漠腹地人工绿地三种灌木的离子吸收特性. 干旱区研究, 19(1): 49–52. |
| [] | Simms E L. 2000. Defining tolerance as a norm of reaction. Evolutionary Ecology, 14: 563–570. DOI:10.1023/A:1010956716539 |