 2012, Vol. 48
2012, Vol. 48文章信息
- 刘振, 李红丽, 董智, 李钢铁, 万玲玲, 岳永杰
- Liu Zhen, Li Hongli, Dong Zhi, Li Gangtie, Wan Lingling, Yue Yongjie
- 浑善达克沙地2种生境下榆树种群空间点格局
- The Spatial Point Pattern of Ulmus pumila Population in Two Habitats in the Otindag Sandy Land
- 林业科学, 2012, 48(1): 29-34.
- Scientia Silvae Sinicae, 2012, 48(1): 29-34.
-
文章历史
- 收稿日期:2011-02-25
- 修回日期:2011-05-05
-
作者相关文章



2. 内蒙古农业大学林学院 呼和浩特 010019
2. College of Forestry, Inner Mongolia Agricultural University Hohhot 010019
种群分布格局是指种群个体在水平空间的配置或分布状况,反映种群个体在水平空间上彼此间的相互关系,与物种的生物学特性、种间竞争以及生境条件等密切相关(Dale,1999)。植物种群的空间格局不仅因物种而异,而且同一物种在不同发育阶段、不同的生境条件下也有明显差别(江洪,1992)。格局研究不仅可对种群的水平结构给以定量的描述,还可用以揭示种群的动态变化,一直是生态学的研究热点(张金屯等,2004; 李先琨等,2002)。传统的格局分析方法只能分析一种尺度下的分布格局,得出的结果与实际的群落结构有较大偏差,难以全面反映种群的分布类型与空间尺度的关系,而点格局分析法在很大程度上克服了传统分析法的不足(张金屯等,2004; 杨洪晓等,2006; Ripley,1977; 1981)。点格局分析法以植物个体构成的空间分布的点图为基础进行格局分析(Ripley,1981; Dale et al., 2001; 张金屯,1998),可以分析各种尺度下的种群格局和种间关系,在拟合分析的过程中最大限度地利用坐标图的信息,因而检验能力较强(张金屯,1998)。自20世纪90年代以来,我国学者运用点格局分析法相继开展了不同乔木树种种群的空间分布格局与空间关联性研究,从不同尺度揭示了种群的分布格局与形成,森林的建立、生长、死亡和更新等生态过程(雷相东等,2002; 汤孟平等,2004; 汤孟平,2010; 张金屯等,2004; 牛丽丽等,2008; 王磊等,2010)。
榆树(Ulmus pumila)疏林广泛分布于浑善达克沙地,是适应半干旱草原气候、并在沙生演替系列中形成的顶极性群落,尤其是在锡林郭勒盟正蓝旗的榆树疏林,其地带性顶极群落表现得更为明显(中国科学院内蒙古宁夏考察队,1985)。榆树根系发达,蒸腾强度小,冠幅大,空间利用充分,具有喜光、耐寒、耐旱、抗病性强等特点,是沙地中重要的保土、防风、固沙树种(刘建等,2003; 马毓泉,1990; 李永庚等,2003)。目前,一些学者对浑善达克沙地榆树疏林生态系统的空间异质性、根系生物量、人为干扰影响及种群大小结构与格局等进行了研究(刘建等,2003; 2005;李永庚等,2003; 赵丽等,2009; 李红丽等,2002),但对不同生境条件下、不同发育阶段榆树种群格局的研究还未见报道。为此,本文采用典型大样地法,调查了固定沙丘和丘间低地2种生境条件下榆树疏林种群的结构,并利用点格局分析法对种群在不同生长阶段的空间分布格局及空间关联性进行分析,以期为沙地榆树疏林的保护和可持续利用提供理论依据。
1 研究区概况研究区位于浑善达克沙地南缘正蓝旗境内桑根达来(42°42'05″ N,115°58'10″ E),该区属温带半干旱区大陆性气候,年均气温1.8 ℃,极端最低温度-38 ℃,≥ 10 ℃年积温2 000 ℃; 年均降水量313.8 mm,降水量年内分配不均,其中夏季占68.3%。研究区中固定沙丘、半固定沙丘、流动沙丘和丘间低地镶嵌分布; 榆树种群(单株散生或以疏林形式分布)多呈丛状分布,主要集中在固定沙丘和丘间低地。固定沙丘上植物种较为丰富,主要有褐沙蒿(Artemisia intramongolica)、糙隐子草(Cleistogenes squarrosa)、沙生冰草(Agropyron desertorum)、苔草(Carex duriuscula)、叉分蓼(Polygonum divaricatum)、防风(Saposhnikovia divaricata)、黄柳(Salix gordejevii)、楔叶茶藨子(Ribes diacanthum)等,土壤为固定风沙土。丘间低地植物茂密,形成草甸植被。优势种为耧斗叶绣线菊(Spiraea aquilegifolia)、小红柳(Salix microstachya var. bordensis)、砂杞柳(Salix kochiana)、羊草(Aneurolepidium chinense)、轮叶沙参(Adenophora tetraphylla)、地榆(Sanguisorda officinalis)、蒲公英(Iaraxacum falciobums)、芦苇(Phragmites australis)等,土壤为草甸土、盐化草甸土。
2 研究方法 2.1 样地设置与调查2009年8月,在全面踏查的基础上,以浑善达克沙地中部正蓝旗桑根达来境内生长的天然沙地榆树种群为研究对象,分别在丘间低地(A)及固定沙丘(B)选取封育的榆树疏林,建立100 m×100 m的典型样地,记录样方内胸径≥2.5 cm榆树的空间位置,量测其胸径、树高、冠幅和枝下高,并记录样地内天然更新的榆树幼苗数量。其中丘间低地为2000年封育,密度为130株·hm-2,固定沙丘榆树为2003年封育,密度为147株·hm-2。
2.2 空间格局分析由于实际测得树木的年龄比较困难,所以采用空间代替时间的方法即径级代替龄级的方法(周晓峰等,2002)。基于此,根据彭羽(2005)对浑善达克沙地榆树标准木的伐桩年轮提出的拟合方程,确定被调查榆树的年龄并分级(表 1)。
|
|
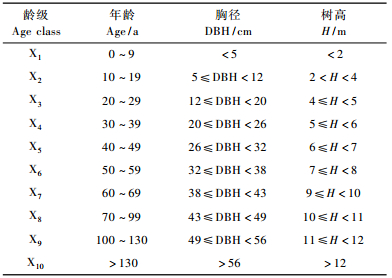
为简化不同龄级立木的格局分析,将榆树种群分为3个发育阶段,即幼树、中龄树和成熟树。利用龄级的划分标准并结合此次调查的实际情况,将X1龄级定为幼树(Ⅰ),X2~X3定为中龄树(Ⅱ),X4~X10定为成熟树(Ⅲ)。利用ADE-4软件对不同发育阶段的榆树种群空间分布格局及其空间关联性进行分析。
2.2.1 种群不同发育阶段个体点格局分析运用Ripley's K点格局法进行榆树空间分布格局分析,其基本公式为:
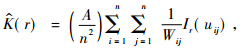
|
(1) |
式中: A为样地面积; n为总点数; Ir(uij)为指示函数,uij为点i和点j之间的距离,当uij ≤ r时,Ir(uij)=1,当uij>r时,Ir(uij)=0;Wij为权重值,表示以点i为圆心、uij为半径的圆周长在面积A中的比例。
为了更直观地解释实际的空间格局,Besag(1977)提出了Ripley的L函数,其公式为: 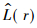
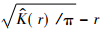
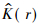
利用ADE-4软件通过10 000次Monte Carlo随机模拟计算99%的置信区间,当
2个发育阶段的空间关联性分析实际上是2个发育阶段种群的点格局分析,其定义和计算原理与单种格局相近(Ripley,1977; Dale et al., 2001),只是将式(1) 中的总点数以种1和种2的个体数n1和n2代之,i和j分别代表种1和种2的个体。同样,可求得Ripley的L函数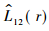
丘间低地和固定沙丘榆树种群不同发育阶段空间分布格局的分析结果见图 1。丘间低地中,榆树幼树(A Ⅰ)在0~28 m尺度上呈聚集分布,在>28 m尺度上呈现随机分布; 在r=5 m时达到最大聚集强度17.65,聚集规模为78.5 m2。中龄树(A Ⅱ)在0~14 m尺度上呈聚集分布,在>14 m尺度上呈现随机分布; 在r=6 m时达到最大聚集强度2.78。成熟树(AⅢ)在0~10 m,27~50 m尺度上呈聚集分布,在10~27 m呈随机分布。由此,丘间低地环境中,榆树种群整体在<10 m的小尺度上呈现聚集分布,幼树和中龄树在大尺度上呈随机分布,而成熟树呈现聚集分布。

|
图 1 丘间低地和固定沙丘榆树种群3个发育阶段的点格局 Figure 1 Point pattern analysis for three developmental stages of U. pumila population in sandy lands between dunes and on fixed sand dunes 实线表示由实际数据计算所得的值,虚线表示用Monte Carlo检验的置信区间(置信水平为99%)。Solid line indicated value of the summary function for the data point pattern,broken line indicated confidence interval with Monte Carlo test.the number of simulated point pattern was 99%.下同The same below. |
固定沙丘中,榆树幼树(B Ⅰ)整体呈聚集分布,在r=35 m时达到最大聚集强度。中龄树(BⅡ)在0~10 m尺度上呈聚集分布,在r=3 m,
2种生境条件下榆树种群的不同发育阶段整体上均在小尺度呈现集群分布,表现出明显的“小聚集、大分散”特点,形成现在沙地最为多见的簇丛状分布格局。对比2种生境同一龄级榆树种群的分布格局,固定沙丘中的榆树种群在达到最大聚集强度时的聚集半径均小于在丘间低地的聚集半径,尤其固定沙丘幼树整体全部为聚集分布,而丘间低地则在0~28 m间为聚集分布,这说明,即使同一龄级的个体,因环境的异质性导致不同尺度下的分布格局存在明显的差别。
3.2 榆树种群不同发育阶段空间关联性丘间低地和固定沙丘榆树种群不同发育阶段空间关联如图 2。丘间低地种群中,幼树(AⅠ)与中龄树(AⅡ)、幼树(AⅠ)与成熟树(AⅢ)分别在r为0~6 m和r为0~4 m时,

|
图 2 丘间低地和固定沙丘榆树种群任意2个发育阶段空间关联 Figure 2 Spatial association of two developmental stages of U. pumila population in sandy lands between dunes and on fixed sand dunes |
固定沙丘种群中,幼树(B Ⅰ)与中龄树(BⅡ)、幼树(BⅠ)与成熟树(BⅢ)在整个分析尺度上的空间关联性都很小; 中龄树与成熟树在2 ~5 m尺度上,呈现空间正关联,在其他尺度上,关联性很小。
2种生境下榆树幼树与中龄树和成熟树的空间关联性表现不同,这极有可能是生境条件造成的; 而中龄树与成熟树间均在小尺度上存在显著正关联,大尺度上无关联性。这种空间关联性的分布格局与种群的空间分布格局相一致。由此推断,榆树种群在较小的空间尺度上倾向于非随机分布,具有明显的空间关联; 在大于某临界尺度时倾向于随机分布,空间关联也变得微弱。
4 结论与讨论种群分布格局是植物与环境长期相互作用的结果(王巍等,1999)。树种在不同发育阶段、不同龄级会表现出不同的空间格局,这与其自然稀疏过程、干扰格局以及环境的变化有密切关系(Greig-Smith,1983)。一般来说,物种的幼龄个体主要呈聚集分布,成年个体表现为随机或均匀分布(张健等,2007; Laaka-Lindberg et al., 2000)。幼龄个体呈现聚集分布主要取决于树种的繁殖特性及种子传播方式(Laaka-Lindberg et al., 2000; Hedenås et al., 2000; 2003),而成年个体的随机或均匀分布与种内及种间竞争、环境的影响有关。本研究中,榆树幼树的群集性生长的特征形成聚集分布,而这种聚集状态在小尺度上延续到榆树整体的发育阶段,尤其是固定沙丘中的榆树种群表现最为明显。固定沙丘的榆树种群分布格局随着龄级的增长,最大聚集强度和聚集半径均减小。同时,固定沙丘中龄树和成熟树种群在达到最大聚集强度时的聚集规模分别为28.26 m2和12.56 m2,均小于在丘间低地的种群。这与2种生境下种子散布机制、风沙与水分的差异有密切关系。就种子散布而言,沙地榆树种子成熟后,借助于风力散布,在母树周围、风力减弱区、灌草丛、甚至牲畜蹄迹中种子呈聚集分布,可导致幼苗与幼树呈聚集分布。固定沙丘地形起伏,风速随地形变化明显,由迎风坡脚向丘顶增大并在背风坡进一步降低,凭借风力扩散的榆树种子在迎风坡脚、背风坡及植被丰富地段等风速减弱区易聚积在一起,形成栖息岛效应,导致幼苗的聚集分布。而且,榆树小尺度的集群生长可以相互减小风蚀的影响,适应沙丘上恶劣的风沙环境,有利于个体生存和种群的更新(赵丽等,2009)。此外,固定沙丘的榆树中龄树和成熟树种群还存在成对现象,这也与二者在小尺度上呈空间正关联相互印证。榆树个体与其邻体形成一定的“小聚集”斑块,这些斑块随机镶嵌分布在沙丘上,使得固定沙丘的榆树中龄树和成熟树种群在大尺度上呈现随机分布,并表现出“大分散”的特点。丘间低地中龄树和成熟树的聚集分布尺度均大于固定沙丘,这与丘间低地地下水位较高,土壤水分较好密切相关。丘间低地地势平坦,风速均匀,在大尺度上整体上呈现随机分布,但受地表植被削弱风速效应的影响而在植被密集处集聚,因而在0~28m的尺度上均呈聚集分布。成熟树在大尺度上呈现聚集分布,则可能是由于丘间低地下水位高、水分条件好造成的。显然,沙地榆树空间分布格局是榆树种群适应严酷环境的自我调节方式,但其分布格局的形成和环境的异质性之间作用机制还需进一步研究。
种内联结用来表示同一物种不同龄级在空间分布上的相互关联性,是对一定时期内植物种群组成个体之间相互关系的静态描述(王磊等,2010)。同一树种不同龄级间的关系主要取决于环境的均匀性、受干扰程度和历史因素等(Leathwick et al., 1992)。本研究中,丘间低地幼树种群的分布在小尺度上与中龄树和成熟树分布均呈空间负关联,幼树的分布受到中龄树和成熟树的排斥; 但在固定沙丘中呈现无关联性。原因可能为丘间低地生境下,中龄树与成熟树的密度高,冠幅大,郁闭度高,林窗小,而榆树有喜光的特性,幼树只有林窗中才能生长。固定沙丘由于地形存在高差,且榆树的郁闭度较低,林下光照条件良好,不会影响幼树的生长。这说明,在郁闭度高的沙地榆树种群中,林冠空隙可能是推进种群更新的积极因素,这与其他学者的研究结果一致(张金屯等,2004)。2种生境中的中龄树与成熟树均在小尺度上存在显著正关联,这与它们在小尺度上的聚集分布具有一致性,说明它们之间的竞争关系很小,对环境的要求和适应性具有一致性。
浑善达克沙地榆树疏林草地是浑善达克沙地植被演替的顶级群落,也是最适合当地气候条件的生态群落。种群聚集强度最大时的聚集规模,可能是种群自然更新的最适面积(Brokaw,1987)。应遵循沙地榆种群分布格局规律和种内联结关系,尽量维持种群稳定和促进种群发展,并根据生境条件及种内联结的关系进行保护和抚育管理,减少树种间对水分及养分的竞争,实现其有序更新。而且,在进行榆树人工林建设时,可依据榆树天然的空间分布格局特征,使造林密度符合天然榆树疏林种群的分布格局及其最大聚集面积尺度,进而应用近自然林业的经营方式进行抚育管理。即在人工造林初期,进行适当密植,使幼苗形成聚集格局,并进行封育、禁牧; 从封育的第4年开始选择生长健壮的幼苗进行定苗。在固定沙丘中,一般以3~5棵为一丛进行定苗; 在丘间低地对单株幼苗进行定苗,并对留存的榆树幼树采用除草、松土等抚育措施。在榆树进入中龄林后,进行适当的择伐和林下补植,促进榆树生长及更新,营造异龄林。榆树达到成熟阶段后,要采取适度的透光抚育,增大林冠空隙,促进林下幼苗及幼树的天然更新。
江洪. 1992. 云杉种群生态学[M]. 北京: 中国林业出版社: 23-98.
|
雷相东, 唐守正. 2002. 林分结构多样性指标研究综述[J]. 林业科学, 38(3): 140-146. DOI:10.11707/j.1001-7488.20020325 |
李红丽, 董智, 王林和, 等. 2002. 浑善达克沙地榆树根系分布特征及生物量研究[J]. 干旱区资源与环境, 16(4): 99-105. |
李先琨, 苏宗明, 向悟生, 等. 2002. 濒危植物元宝山冷杉种群结构与分布格局[J]. 生态学报, 22(12): 2246-2253. DOI:10.3321/j.issn:1000-0933.2002.12.031 |
李永庚, 蒋高明, 高雷明, 等. 2003. 干扰对浑善达克沙地榆树疏林的影响[J]. 植物生态学报, 27(6): 829-834. |
刘建, 刘凤红, 董鸣, 等. 2005. 浑善达克沙地南缘榆树种群的大小结构和邻体格局[J]. 中国沙漠, 25(1): 75-80. |
刘建, 朱选伟, 于飞海, 等. 2003. 浑善达克沙地榆树疏林生态系统的空间异质性[J]. 环境科学, 23(4): 29-34. |
马毓泉. 1990. 内蒙古植物志(第2卷, 2版): 呼和浩特: 内蒙古人民出版社, 105-115. http://www.oalib.com/paper/4831141
|
牛丽丽, 余新晓, 岳永杰. 2008. 北京松山自然保护区天然油松林不同龄级立木的空间点格局[J]. 应用生态学报, 19(7): 1414-1418. |
彭羽. 2005. 浑善达克沙地退化生态系统生态恢复的自然保护区途径[J]. 中国科学院植物研究所博士学位论文. |
汤孟平. 2010. 森林空间结构研究现状与发展趋势[J]. 林业科学, 46(1): 117-122. DOI:10.11707/j.1001-7488.20100119 |
汤孟平, 唐守正, 雷相东, 等. 2004. 林分择伐空间结构优化模型研究[J]. 林业科学, 40(5): 25-31. DOI:10.11707/j.1001-7488.20040504 |
王磊, 孙启武, 郝朝运, 等. 2010. 皖南山区南方红豆杉种群不同龄级立木的点格局分析[J]. 应用生态学报, 21(2): 272-278. |
王巍, 刘灿然, 马克平, 等. 1999. 东灵山两个落叶阔叶林中辽东栎种群结构和动态[J]. 植物学报, 41(4): 425-432. |
王峥峰, 安树青, 朱学雷, 等. 1998. 热带森林乔木种群分布格局及其研究方法的比较[J]. 应用生态学报, 9(6): 575-580. |
杨洪晓, 张金屯, 吴波. 2006. 毛乌素沙地油蒿种群点格局分析[J]. 植物生态学报, 30(4): 563-570. |
张健, 郝占庆, 宋波, 等. 2007. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J]. 应用生态学报, 18(8): 1681-1687. |
张金屯. 1998. 植物种群空间分布的点格局分析[J]. 植物生态学报, 22(4): 344-349. |
张金屯, 孟东平. 2004. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报, 24(1): 35-40. |
赵丽, 王晓江, 刘果厚, 等. 2009. 浑善达克沙地榆树种群结构、格局及动态研究[J]. 中国沙漠, 29(3): 508-513. |
中国科学院内蒙古宁夏考察队. 1985. 内蒙古植被[M]. 北京: 科学出版社: 785-789.
|
周晓峰, 王晓春, 韩士杰, 等. 2002. 长白山岳桦苔原过渡带动态与气候变化[J]. 地学前缘, 9(1): 227-231. |
Besag J E. 1977. Comments on Ripley's paper[J]. Journal of the Royal Statistical Society: Series B(Methodological), 39(2): 193-195. |
Brokaw N V L. 1987. Gap phase regeneration of three pioneer tree species in a tropical forest[J]. Journal of Ecology, 75(1): 9-20. DOI:10.2307/2260533 |
Dale M R T. 1999. Spatial pattern analysis in plant ecology[M]. London: Cambridge University Press: 207-276.
|
Dale M R T, Powell R D. 2001. A new method for characterizing point patterns in plant ecology[J]. Journal of Vegetation Science, 12(5): 597-608. DOI:10.2307/3236899 |
Greig-Smith P. 1983. Quantitative plant ecology[M]. London: Blackwell Scientific Publicanons.
|
Hedenås H, Bolyukh V O, Jonsson B G. 2003. Spatial distribution of epiphytes on Populus tremula in relation to dispersal mode[J]. Journal of Vegetation Sciene, 14(2): 233-242. |
Hedenås H, Ericson L. 2000. Epiphytic macrolichens as conservation indicators: Successional sequence in Populus tremula stands[J]. Biological Conservation, 93(1): 43-53. DOI:10.1016/S0006-3207(99)00113-5 |
Ripley B D. 1977. Modelling spatial pattern[J]. Journal of the Royal Statistical Society: Series B(Methodological), 39(2): 172-212. |
Ripley B D. 1981. Spatial statistics[M]. New York: John Wiley and Sons: 10-200.
|
Laaka-Lindberg S, Hedderson T A, Longton R E. 2000. Rarity and reproductive characters in the British hepatic flora[J]. Lindbergia, 25(2): 78-84. |
Leathwick J R, Mitchell N D. 1992. Forest pattern, climate and volcanism in central North Island, New Zealand[J]. Journal of Vegetation Science, 3(5): 603-616. DOI:10.2307/3235827 |
Sotirios K, George A B. 2005. Spatial relationships between tree species and gap characteristics in broad-leaved deciduous woodland[J]. Journal of Vegetation Science, 16(5): 587-596. DOI:10.1111/j.1654-1103.2005.tb02400.x |