 2011, Vol. 47
2011, Vol. 47文章信息
- 姜俊, 赵秀海
- Jiang Jun, Zhao Xiuhai
- 吉林蛟河针阔混交林群落优势种群种间联结性
- Interspecific Correlations among Dominant Tree Species in the Coniferous and Broad-Leaved Mixed Forest Communities in Jiaohe, Jilin Province
- 林业科学, 2011, 47(12): 149-153.
- Scientia Silvae Sinicae, 2011, 47(12): 149-153.
-
文章历史
- 收稿日期:2011-06-08
- 修回日期:2011-11-02
-
作者相关文章



种间联结性是指不同物种在空间分布上的相互关联性,是植物群落重要的数量和结构特征之一(邓贤兰等, 2003),是群落形成、维持和演替的基础(王文进等, 2007)。种间联结能有效量化群落中物种相互作用和物种在环境中的配置情况(周先叶等, 2000),对于正确认识群落的结构组成、功能和群落演替动态具有重要意义,能为森林经营、森林植被恢复和生物多样性保护提供理论依据(张金屯等, 2003)。当前对群落种间关联性的研究主要集中在2方面:有无关联和关联程度。形成种间联结性的因素有很多,国内外研究认为种间联结性与下列4个因素有关:1)相似与不相似的环境需求;2)一个种为另外一个种创造了定居条件或者前者对后者施加了压力;3)2者在资源竞争中互相排斥;4)2者通过物理或化学作用互相影响(Greig-Smith, 1983;Kershaw et al., 1985;周先叶等, 2000)。
长期以来对针阔混交林的不合理经营和人为干扰造成群落物种多样性逐渐降低、目的树种丧失严重, 如何恢复和改造针阔混交林成为亟待解决的问题,这需要了解和掌握群落中各个优势树种之间的相互关系。本研究通过χ2检验、Pearson相关系数检验和Spearman秩相关系数检验分析针阔混交林内自然更新的主要优势乔木种间关联性,并结合植物种群对环境主导因子的不同生态适应性来划分优势树种的生态种组,为该区域针阔混交林生态恢复和林木抚育及保护提供科学依据。
1 研究区概况研究区位于吉林省蛟河林业试验区管理局林场(127°44.111′—127° 44.667′E, 43°57.524′—43°58.042′N),海拔459~517 m,全区属长白山系张广才岭山脉,地势由西北向东南倾斜,北部高,东南平坦开阔。土壤为山地暗棕色森林土壤,土层厚20~100 cm,土壤形态有层次分化,富含有机质。该区域属于受季风影响的温带大陆性山地气候,年均气温3.8 ℃,最热月7月日平均气温21.7 ℃,最冷月1月日平均气温-18.6 ℃。年均降水量700~800 mm。该区植被区系分类属于典型的北温带植物区系的针阔混交林,植物种类多样、分布复杂,森林群落垂直成层现象较为明显。乔木树种主要包括红松(Pinus koraiensis)、沙松(Abies holophylla)、千金榆(Carpinus cordata)、春榆(Ulmus japonica)、色木槭(Acer mono)、簇毛槭(Acer barbinerve)、胡桃楸(Juglans mandshurica)、水曲柳(Fraxinus mandshurica)、紫椴(Tilia amurensis)、白桦(Betula platyphylla)、白牛槭(Acer mandshuricum)、蒙古栎(Quercus mongolica)、大青杨(Populus ussuriensis)、裂叶榆(Ulmus laciniata)和怀槐(Maackia amurensis)等,灌木主要包括暴马丁香(Syringa reticulata)、毛榛(Corylus mandshurica)、瘤枝卫矛(Euonymus pauciflorus)和东北鼠李(Rhamnus davurica)等。
2 研究方法 2.1 采样设计及数据预处理在42 hm2针阔混交林固定监测样地内采用典型取样方法,随机设置100块20 m×20 m样地。调查记录样地内所有胸径>1 cm的植株个体种名、胸径、树高、枝下高及冠幅(东西冠幅长、南北冠幅长),同时记录每块样地的地理坐标和海拔。统计样地里出现的物种和物种个体数,计算出频度、密度以及种对出现的样地数,计算样地内物种的重要值得到样地优势物种,选取重要值前18位的物种(表 1)来计算和分析种间联结性,并结合物种的生态学特性划分出相应的生态种组。
|
|
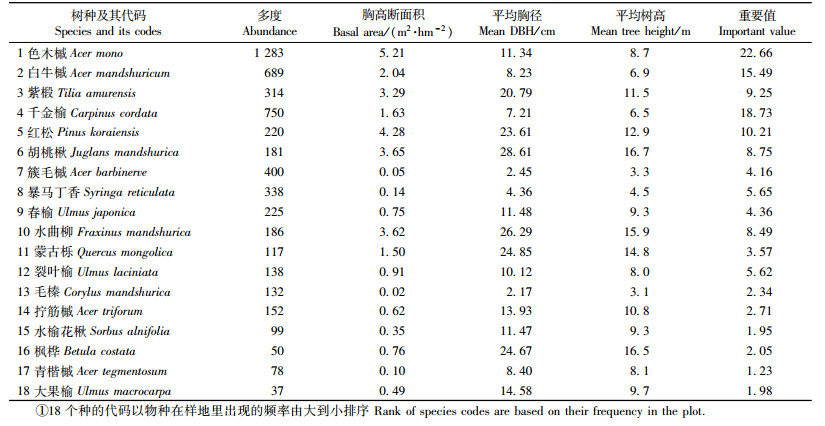
本研究对物种间的总体相关性检验采用方差比率法(Schulter, 1984),按照以下公式计算检验统计量(杜道林等,1995)。
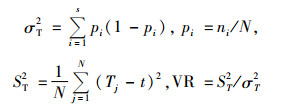
|
N为总样地数;ni为物种i出现的样地数;Tj为研究样地j内出现的物种总数;t为样地内物种的平均数;σT2是总样地数方差;ST2是总物种数方差;VR为全部物种的关联指数,在零假设独立性的条件下,VR的期望值为1,当VR>1时,表明物种间呈现净的正关联;VR < 1时,表明物种间存在净的负关联;VR=1,表明物种间无关联。通过计算统计量W=VR×N来检验物种联结的显著性,在独立假设条件下,若种间联结不显著,则W落在χ0.952(N)≤W≤χ0.052(N)区间内的概率就为90% (Schulter, 1984)。
2.3 种间联结性分析将样地数据建立2×2列联表,根据出现或不出现数据的方差比例,构建18个优势物种在100个样地内的18×100原始数据矩阵,并计算观测值a, b, c和d,其中a为2个物种都出现的样地数,b和c为仅有1个物种出现的样地数,d为2个物种都不出现的样地数(王伯荪等, 1985)。χ2统计量用来检验种对间的联结性,由于取样的非连续性,采用Yates的连续性校正公式计算χ2值(张金屯, 2004)。
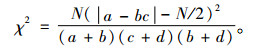
|
当χ2 < 3.841时,认为2个物种独立分布,种间联结独立;χ2>6.635则认为种间联结极显著;3.841 < χ2 < 6.635认为种间联结显著。因为χ2没有负值,种间的正负联结根据ad与bc值来判断,当ad>bc时为正联结,ad < bc为负联结。
2.4 种间相关性测定应用Pearson相关系数和Spearman秩相关系数对2个物种间的线性关系做定量分析,分析2个种对间同时出现的可能性(张金屯, 1995)。
Pearson相关系数rp(i, j)运算公式:
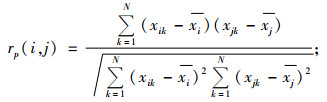
|
Spearman秩相关系数rs(i, j)公式:
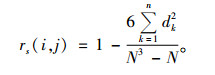
|
dk=(xik-xjk),xik和xjk分别为种i和种j在样地k中的秩。
统计分析、制图均采用R 2.12. 2(R Development Core Team, 2010)统计软件。
3 结果与分析 3.1 总体种间相关性计算总体种间相关性得到方差比率VR=1.206>1,表明在独立零假设条件下,总体种间相关关系表现为净的正联结。计算统计量W检验VR的显著性,W=120.61,查表得出,W落在χ0.952(N)和χ0.052(N)区间之外,说明选择的18种优势物种总体表现为显著的正联结,表明针阔混交林群落结构和物种组成会随着演替的进展趋向于稳定。
3.2 种对间关联性针阔混交林优势种的153个种对里,检验呈正联结的有88对,占总种对数的57.5%,负联结的有65对,占总对数的42.5%,种间呈极显著和显著正联结的23对,占总对数的15%;一般正联结的65对,占总对数的42.5%;极显著和显著负联结的56对,占总对数的36.6%,一般负联结的9对,占总对数的5.9%(图 1)。种对间的相互联结性体现了物种之间的依赖性与排斥性。正联结种对多是阳性或中性偏阳树种,对环境或资源(光照)有相似的需求,存在显著负联结的种对的生态学特征不同,对生境的要求不一样,即呈现相互排斥性。检验中正联结种对数较多,说明种群对生境和资源要求有一定的相似性,与总体种间相关性检验的方差比率VR值的结果一致,群落中优势物种处于稳定阶段。

|
图 1 优势树种种间关联的χ2半矩阵图 Figure 1 Semi-matrix graph of interspecific correction χ2-test of association of dominant tree species ●:极显著正相关Positive correlation at 0.01 level; ▲:显著正相关Positive correlation at 0.05 level;+:不显著正相关Not significant positive correlation; ■:极显著负相关Negative correlation at 0.01 level;◆:显著负相关Negative correlation at 0.05 level;*:不显著负相关Not significant negative correlation.下同The same below. |
张金屯(2004)研究表明,单独使用χ2检验研究种间的联结性存在一定缺陷,需要结合Spearman秩相关系数检验和Pearson相关系数检验做更全面的分析,图 2和3分别为18个优势种的Spearman秩相关系数检验和Pearson相关系数检验的半矩阵图。Pearson相关系数检验中正相关的种对数为67个,占总种对数的43.8%,负关联种对有85个,占总种对数的55.6%。Spearman秩相关系数检验中正联结118对,占总种对数的77.1%,负联结35对,占总种对数的22.9%。2个系数检验的正联结种对数明显大于负联结的种对数,说明18个优势种对间大多呈正联结。Spearman秩相关系数检验的正联结比例要比χ2检验和Pearson相关系数检验高,谢湉等(2010)认为如果2个重要值的秩较小时,用Spearman检验会认定种间关联为正,出现与χ2检验和Pearson相关系数检验不同的结果。

|
图 2 优势树种种间Spearman秩相关系数半矩阵图 Figure 2 Semi-matrix of Spearman's rank correlation coefficients of dominant tree species |

|
图 3 优势树种种间Pearson相关系数半矩阵图 Figure 3 Semi-matrix of Pearson correlation coefficients of dominant tree species |
综合上述相关系数检验结果以及植物种群对环境的不同生态适应性和生物学特征,划分了针阔混交林优势种群的生态种组。原则是以环境适应性相似的物种分为一组,同一生态种组的物种间尽可能多的正联结,不同生态种组的物种为负联结(郭志华等,1997)。大致将18个优势种群分为3个不同的生态种组。
生态种组Ⅰ包括色木槭、紫椴、千金榆、红松、拧筋槭(Acer triforum)和水榆花楸(Sorbus alnifolia)。该组树种之间多呈现正联结,这些树种是乔木层的绝对优势树种或主要伴生种,且属于演替后期的群落主导树种,个体数量多且生态幅广、耐荫,其中色木槭和紫椴是针阔混交林主要的软阔叶树种,红松是针阔混交林森林经营的最重要的目的树种。
生态种组Ⅱ包括簇毛槭、暴马丁香、毛榛、枫桦(Betula costata)和大果榆(Ulmus macrocarpa)。在种组内种对联结程度不大,每个物种的独立性较强。该组主要以林下灌木为主,在针阔混交林群落分布较广,多属中性或中性偏阳的植物,对阳光的需求不强,具有一定的耐荫性,常见于沟谷或受干扰较严重的地方。
生态种组Ⅲ包括白牛槭、胡桃楸、春榆、水曲柳、蒙古栎、裂叶榆和青楷槭(Acer tegmentosum)。该组内种间多表现为负联结,树种大多为喜阳耐旱的乔木,生长在阳坡、半阳坡,土壤湿度较低且光照较好的生境内,分布较为集中,大多数是演替初期的先锋种,与种组Ⅰ物种,尤其是乔木层的优势种多成负联结。
4 结论与讨论针阔混交林样地群落的种间联结结果表明,18种优势种总体联结性呈正相关,多数种对间正联结程度较松散,基本上趋于独立出现,有些种群间出现显著的负联结,整体呈近似钟形的正态分布曲线,大部分无显著关联的围绕中点,其他种间关系分别处于曲线的两端(显著的正联结和显著负联结),可能与当前群落演替阶段及物种本身的生态学特性有关。周先叶等(2000)认为正负关联比越高, 群落结构越趋于稳定, 多物种可以稳定共存, 群落达到与环境条件相适应的稳定阶段。结合物种组成、年龄结构和重要值特征等,可以认为该森林群落处于先锋树种衰退, 红松、色木槭为主的多个优势树种共存的林分发育阶段,随演替的进展主要树种间逐渐会出现显著的正联结或负联结,群落朝着有利于物种稳定和共存的方向发展。
史作民等(2001)研究发现,物种间的联结性与其生态位有很大相关性,种间联结性强的种对,一般有高的生态位重叠,种间联结性低的种对会出现生态位分化。本研究的针阔混交林群落大多数种对呈正联结体现了物种间生态位重叠。种对负联结反映了物种生态位的分离,如红松是演替后期的耐荫性树种,白牛槭是喜光阳性树种,种对表现为负联结。
本研究生态种组的划分可为营林提供依据,如在设计针阔混交林造林方案时,在种组内选择搭配树种,利于针阔混交林林下植被的稳定和发展。
邓贤兰, 刘玉成, 吴杨. 2003. 井冈山自然保护区拷属群落优势种群的种间联结关系研究[J]. 植物生态学报, 27(4): 531-536. DOI:10.17521/cjpe.2003.0077 |
杜道林, 刘玉成, 李睿. 1995. 缙云山亚热带栲树林优势种群间联结性研究[J]. 植物生态学报, 19(2): 215-220. |
史作民, 刘世荣, 程瑞梅, 等. 2001. 宝天曼落叶阔叶林种间联结性研究[J]. 林业科学, 37(2): 29-35. DOI:10.11707/j.1001-7488.20010204 |
王伯荪, 彭少麟. 1985. 南亚热带常绿阔叶林种间联结测定技术研究[J]. I.种间联结测式的探讨与修订.植物生态学与地植物学丛刊, 9(4): 274-285. |
王文进, 张明, 刘福德, 等. 2007. 海南岛吊罗山热带山地雨林两个演替阶段的种间联结性[J]. 生物多样性, 15(3): 257-263. |
谢湉, 巨天珍, 师贺雄, 等. 2010. 甘肃省小陇山濒危珍稀植物白皮松群落种间关联[J]. 生态学杂志, 29(3): 448-453. |
张金屯. 1995. 数量生态学[M]. 北京: 科学出版社.
|
张金屯. 2004. 植被数量生态学方法[M]. 北京: 中国科学技术出版社..
|
张金屯, 焦蓉. 2003. 关帝山神尾沟森林群落木本植物种间联结性与相关性研究[J]. 植物研究, 23: 458-463. DOI:10.3969/j.issn.1673-5102.2003.04.020 |
周先叶, 王伯荪, 李鸣光, 等. 2000. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析[J]. 植物生态学报, 24(3): 332-339. |
Greig-Smith P. 1983. Quantitative plant ecology[M]. Oxford: Blackwell Science Publications: 105-112.
|
Kershaw K A, Looney J H. 1985. Quantitative and dynamic plant ecology[M]. London: Edward Arnold Limited: 78-94.
|
Schluter D. 1984. Avariance test for detecting species association with some example applications[J]. Ecology, 65(3): 998-1005. DOI:10.2307/1938071 |