 2011, Vol. 47
2011, Vol. 47文章信息
- 卢军, 李凤日, 张会儒, 张守攻
- Lu Jun, Li Fengri, Zhang Huiru, Zhang Shougong
- 帽儿山天然次生林主要阔叶树种叶量分布模拟
- Simulation of Foliage Distribution for Major Broad-Leaved Species in Secondary Forest in Mao'er Mountain
- 林业科学, 2011, 47(12): 114-120.
- Scientia Silvae Sinicae, 2011, 47(12): 114-120.
-
文章历史
- 收稿日期:2010-03-15
- 修回日期:2010-10-27
-
作者相关文章


2. 东北林业大学林学院 哈尔滨 150040;
3. 中国林业科学研究院林业研究所 北京 100091
2. Forestry School, Northeast Forestry University Harbin 150040;
3. Research Institute of Forestry, CAF Beijing100091
叶量的空间分布是树冠结构的基本组成部分,它直接影响树冠内光能的分布,进而影响树木的生长。评估叶量的空间分布以及探讨叶量与林木和林分因子的关系能更好地理解通过树冠将能量转化成地上生物量的机制。
近十几年,国内外对枝叶生物量空间分布做了大量研究。Mäkelä等(2001)在研究欧洲赤松(Pinus sylvestris)时发现,如果把树冠分成上、下2部分,在上半部分,叶量与枝条基径处横断面积的比率从梢头向下是逐渐增加的; 而在下半部分,该比率则迅速减小。Xu等(1998)对花旗松(Pseudotsuga menziesii)叶量的水平分布和垂直分布研究表明:单个枝条的叶量随着枝条的基径和相对高度增加而增加,进一步阐述了树木的优势度、树冠大小和林分因子影响着叶量空间分布。Menalled等(2001)通过测定幼树的叶的分布、树冠形状、生物量分配模式,探讨了3种不同类型的树冠结构与生物量的分配策略。Kantola等(2004)应用3个不同年龄组的29株样木,采用修正的冠形模型来描述树干和树冠的动态,表明对于中龄林和成熟林林分叶量密度最大值出现在相对冠高50%~70%处,而对于幼龄林则出现在冠基部。Oliver等(1990)认为:在树冠范围内,叶量(或叶面积)及树干断面积生长量随着着枝深度的增加而不断增加,且二者呈线性关系,而在树冠以下至树干基部不同位置上的树干断面积生长无明显差异。Zhang等(2004)基于树冠内叶量分布和管道模型理论,对集约经营的花旗松人工林枝叶生物量模型进行了预估,认为冠基部树干带皮直径、树冠横断面高和冠长是预估叶量的关键因子。许景伟等(2007)对胶南沙质海岸海防林9年生火炬松(Pinus taeda)幼树的生物量进行研究,也得出了同样的结论。Wang等(1990)确定了树冠大小和外形,针叶树树冠的叶量分布强烈地影响辐射吸收、光合作用和蒸腾作用。在树冠内,叶量水平分布和叶量垂直分布都不是均衡的(Assmann,1970),枝条水平上的叶量模型(Burk et al., 1983; Maguire et al., 1996)可以有效地组合在现有的冠形模型与枝条分布模型中来估计叶面积密度的垂直变化趋势,其他的描述枝条叶量水平分布的模型(Kershaw et al., 1996)考虑更详细的二维或三维的叶面积密度的描述。因此,冠形模型通过引入对叶量水平和垂直分布的模拟,对描述树木结构起到重要的作用。
以往对于叶量分布的研究都是针对针叶树种的,而对于阔叶树种的叶量分布研究很少,而且以往的研究大多采用各种分布函数对枝叶生物量的空间分布进行拟合,对不同林分条件下阔叶树种树木叶量或叶面积在树冠内的空间分布格局的差异、枝条着叶量和枝条生长的关系的研究比较少。本研究针对天然次生林内阔叶树种的叶量规律,建立累积叶量分布模型,这对于树木叶量的研究具有探索性意义。
1 研究地概况本研究选择具有典型黑龙江省森林特点的东北林业大学帽儿山试验林场为研究地。帽儿山试验林场位于黑龙江省东南部,地理坐标为127°30′—127°34′E,45°20′—45°25′N。总面积为26 496 hm2,南北长30 km,东西宽20 km,共151个林班。该地区属温带季风气候,但是具明显的大陆性气候特征,四季分明,冬长夏短。地带性土壤为暗棕壤,有机质、各种化学元素、微量元素含量都较高。植被属于长白山植物区系,是由地带性顶级植被阔叶红松(Pinus koraiensis)林经人为干扰破坏后形成的较典型的东北东部天然次生林。
2 数据收集与整理2007年共设置了30块天然次生林标准地,位于帽儿山试验林场的中林和跃进施业区,所选的标准地所在林分均是未经过间伐、生长正常的次生林,每块标准地都有GPS定位,根据地形和林相特点,设置不同面积的标准地,一般为20 m×50 m,最大为30 m×70 m。
2.1 解析木的选取及测定由于次生林林分状况复杂,树种结构也很复杂,因此在选取解析木时,不能使用传统的等断面积分级法,只能根据计算得出的树种组成和每木检尺的结果来选取标准地外的几个优势树种的平均木和优势木,以此作为标准地内林分主要树种最大的和平均的生长状况。一般每块标准地对应选择2或3种优势树种,然后每个树种选择1株优势木,1或2株平均木。
2.2 枝解析外业测定1) 解析木伐倒后,首先确定第一活枝的高度,即树冠基部处的高度,测定树冠基部到梢头之间的距离,即冠长(CL);
2) 把整个冠长即活树冠部分根据冠长平均分成3部分,从上至下依次编为上层、中层和下层;
3) 从梢头开始,以每个区分段为单位,进行枝解析外业的测定;
4) 为了分析枝条基径和枝长的生长及树冠结构的动态变化,在每一树冠区分段内要选择1个或多个具有平均基径和枝叶生长状态的枝条作为标准枝进行分析。
2.3 叶生物量的测定叶的生物量采用以质量为基准的标准枝法进行。通过野外采样和实验室称重(精度0.000 1 g电子天平)的方法来测定树叶的生物量。
2.4 解析木枝条数据整理在所设置的30块天然次生林标准地中,共获得解析标准木176株,逐株进行树干解析、枝解析及生物量的测定。将野外调查数据按解析样木、解析样枝进行归类、统计,建立数据库。176株解析木总共测量了3 401个活枝条。所有枝条叶量的最后统计结果见表 1。
|
|

常用几种描述单木叶量的分布模型主要有正态分布、β分布和Weibull分布。在这些模型中,Weibull分布由于其具有较大的灵活性和适应性,既能较好地拟合不同偏度、峰度的单峰山状曲线,又能拟合反J型递减曲线。因此,自Smalley等(1973)引用Weibull分布密度函数描述林分直径分布以来,在林业中逐步得到广泛应用,并取得良好的效果。
树冠叶量的垂直分布可以用2参数的Weibull分布函数描述:
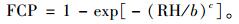
|
(1) |
式中:FCP为叶量累积百分比; RH为相对着枝深度; b,c是模型的参数。
本研究尝试用这种Weibull分布来描述天然次生林10个阔叶树种的叶量分布时发现:这种分布并不适合阔叶树种,出现了严重的左偏;而利用对数正态(log normal)分布来描述叶量的垂直分布时发现效果良好。图 1和表 2为利用对数正态分布描述的白桦叶量垂直分布。

|
图 1 白桦叶量垂直分布 Figure 1 Vertical foliage distribution of Betula platyphylla Chi-Square test=32.829 56, df =4(adjusted), p=0.000 00 |
|
|
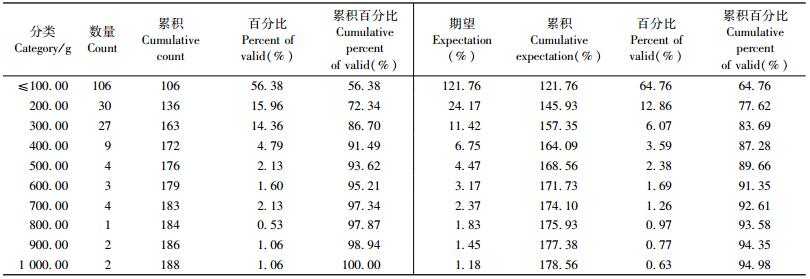
从图 1和表 2中可以看出:绝大多数的叶量都是小于100 g的,其比例超过了50%,小于300 g叶量的累积比例更是超过了86%。因此,用简单的分布形式来描述叶量的垂直分布不能准确地描述树冠内叶量的变化规律。本研究的目标是结合树冠轮廓模型的特点,研究10个树种在不同着枝深度上叶量的垂直分布规律,并建立各树种的叶量分布模型。
3.2 叶量分布模型的设计树木活树冠的累积叶量从树冠基部到梢头的垂直分布服从一个“S”形状(Tahvanainen et al., 2008) (图 2)。为了描述这种叶量的分布,用3参数或者4参数的Sigmoid曲线来拟合。本研究用3参数的Chapman-Richards方程来建立10个阔叶树种的叶量分布模型,所采用的变量是相对高度,而不是绝对高度,这个方程是“S”形曲线。方程的形式如下:

|
图 2 树冠内相对高度上的叶量累积(以白桦为例) Figure 2 Cumulative foliage on relative height in the crown (e.g. Betula platyphylla) |
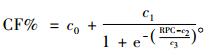
|
(2) |
式中:CF%为树冠内枝条累积叶量百分比; RPC为树冠内枝条的相对高度; c0, c1, c2, c3为模型的参数,c1为尺度参数。
4 结果与分析用最小二乘法来估计公式(2)的参数,得到的结果见表 3。
|
|

参数估计的结果中,在α=0.05的置信度下,相关系数都很高,都超过了0.91,白桦达到了0.98,拟合效果非常好。
做出10个树种的曲线图以及对应的残差分布图,来检验模型拟合的效果(图 3)。

|
图 3 10个树种叶量分布拟合曲线与残差分布 Figure 3 Fitting curves of foliage distribution models and residual distribution for ten species |
这10个树种的残差分布均匀、不发散,说明参数估计结果是无偏的、良好的; 但是这10个树种的曲线形状也不完全相同,水曲柳、榆树和杨树的曲线并不是典型的“S”形曲线,类似于抛物线形,而其他7个树种的曲线都是很明显的“S”形曲线。分析这10个树种累积叶量的分布规律,见表 4。
|
|

对式(2)进行求导即可得出每个树种在冠内相对高度的叶量最大值的位置,并且绘制叶量在冠内相对高度的分布图,见图 4。

|
图 4 冠内相对高度处叶量的分布 Figure 4 Estimated relative accumulation of living crown biomass of ten species |
水曲柳冠内最大叶量出现的位置在0.82 CL处,这是出现的最高位置,而最低的则是黄菠萝,出现在0.52 CL处。其他树种分布在这个区间之内,比如白桦为0.66 CL,柞树为0.78 CL。
每个树种在树冠内相对高度上叶量的垂直分布是不同的,在冠内相对高度0.3 CL处,叶量百分比分布很少,一般都小于0.1,只有黄菠萝是0.115 6;当相对高度达到0.4 CL时,叶量百分比仍然很低,一般都在0.2左右; 当达到0.5 CL,即树冠的一半的位置时,叶量百分比逐渐增大,一般都在0.3左右,黄菠萝达到了0.513 1;随着相对高度的增加,在0.7 CL时这种增加的幅度是最大的,普遍都达到了0.6以上,其中最低的是水曲柳0.520 8,最高的是黄菠萝0.792 9;在0.9 CL时,这种增长的趋势变缓,叶量百分比几乎都达到了0.8~0.9,也就是几乎全树冠的叶量。从这个规律可以看出:天然次生林阔叶树种的叶量在树冠的上部(0.2 CL以下)所占比例很小,枝条都很小,叶量必然就少; 在树冠的中部以及中下部(0.4 CL~0.8 CL)叶量所占比例最大,几乎集中了整个树冠60%~70%的叶量,其中,白桦达到了72.22%,杨树达到了72.55%,水曲柳最低为57.51%,这可能与水曲柳的树叶叶片较大有关系,而在树冠的下部(0.8 CL以下),几乎占很少的叶量,大约10%左右。这些特点也与阔叶树种的生物学特性是相一致的。
5 结论与讨论利用对数正态分布来描述本研究中阔叶树种的叶量垂直分布,取得了较好的效果。建立了10个阔叶树种的单木模型来预测活树冠内叶量的垂直分布,用3参数Chapman-Richards方程来估计叶量在树冠内相对高度处的累积叶量分布,它能很好地适合这10个树种,拟合效果较好。参数估计的结果表明,在α=0.05的置信度下,相关系数很高。其中,尺度参数c1介于0.94~1.46之间,而Tahvanainen等(2008)对芬兰的欧洲赤松、欧洲云杉(Picea abies)和桦木的研究结果中,c1介于0.984~0.993之间。研究中发现,水曲柳冠内最大叶量出现的位置在0.82 CL(冠长)处,这是10个树种中出现的最高位置,而最低的则是黄菠萝,出现在0.52 CL处,其他树种分布在这个区间之内。天然次生林阔叶树种的叶量在树冠的上部(0.3 CL以下)所占比例很小,在树冠的中部以及中下部(0.4 CL~0.8 CL)叶量所占比例最大,几乎集中了整个树冠60%~70%的叶量,其中,白桦达到了72.22%,杨树达到了72.55%,水曲柳最低为57.51%,而在树冠的下部(0.8 CL以下),几乎占很少的叶量,大约10%左右。这与前人的研究都是相类似的,比如,在不同年龄的欧洲赤松林分中,垂直分布的叶量最大值出现在50%相对树高处(Mäkelä et al., 2001),在幼龄的赤松林分中,最大值出现在树冠的中部偏下(Kellomäki et al., 1980)。
本研究所建立的次生林林分内主要树种的叶量分布模型会对不同采伐方案中择伐树木的叶量分布提供无偏的估计,可为过程生长模型及树木三维可视化提供参数; 但是树木种群状态以及林分条件对叶量垂直分布的影响需要进一步研究,也将是未来要开展的工作。
许景伟, 艾淑杰, 韩芳, 等. 2007. 沙质海岸火炬松幼树生物量的研究[J]. 山东农业大学学报:自然科学版, 38(2): 252-256. |
Assmann E. 1970. The principles of forest yield studies[J]. Pergamon Press, Oxford: 506. |
Burk T E, Nelson N D, Isebrands J D. 1983. Crown architecture of short rotation intensively cultured Populus. Ⅲ. A model of first order branch architecture[J]. Can J For Res, 13(6): 1107-1116. DOI:10.1139/x83-148 |
Kantola A, Mäkelä A. 2004. Crown development in Norway spruce[Picea abies(L.)Karst.][J]. Trees-Structure and Function, 18(4): 408-421. |
Kellomäki S, Hari P, Kanninen M, et al. 1980. Eco-physiological studies on young Scots pine stands. II. Distribution of needle biomass and its application in approximating light conditions inside the canopy[J]. Silva Fenn, 14(3): 243-257. |
Kershaw J A, Maguire D A. 1996. Crown structure in western hemlock, Douglas-fir, and grand fir in western Washington: trends in branch-level mass and leaf area[J]. Canadian Journal of Forest Research, 25(12): 1897-1912. |
Maguire D A, Bennett W S. 1996. Patterns in vertical distribution of foliage in young coastal Douglas-fir[J]. Can J For Res, 26(11): 1991-2005. DOI:10.1139/x26-225 |
Mäkelä A, Vanninen P. 2001. Vertical structure of Scots pine crowns in different age and size classes[J]. Trees-Structure and Function, 15(7): 385-392. DOI:10.1007/s004680100118 |
Menalled F D, Kelty M J. 2001. Crown structure and biomass allocation strategies of three juvenile tropical tree species[J]. Plant Ecology, 152(1): 1-11. DOI:10.1023/A:1011495916036 |
Oliver C D, Larson B C. 1990. Forest stand dynamics. McGraw-Hill, Inc[J]. New York, NY: 467. |
Tahvanainen T, Forss E. 2008. Individual tree models for the crown biomass distribution of Scots pine, Norway spruce and birch in Finland[J]. Forest Ecology and Management, 255(3/4): 455-467. |
Wang Y P, Jarvis P G. 1990. Description and validation of an array model——MAESTRO[J]. Agricultural and Forest Meteorology, 51(3/4): 257-280. |
Xu M, Harrington T B. 1998. Foliage biomass distribution of loblolly pine as affected by tree dominance, crown size, and stand characteristics[J]. Can J For Res, 28(6): 887-892. DOI:10.1139/x98-060 |
Zhang Y, Borders B E, Will R E, et al. 2004. A model for foliage and branch biomass prediction for intensively managed fast growing loblolly pine[J]. Forest Science, 50(1): 65-80. |