 2011, Vol. 47
2011, Vol. 47文章信息
- 朱丽华, 应晨希, 叶建仁
- Zhu Lihua, Ying Chenxi, Ye Jianren
- 松材线虫近交系的培育
- Inbreeding of the Pine Wood Nematode, Bursaphelenchus xylophilus
- 林业科学, 2011, 47(12): 92-96.
- Scientia Silvae Sinicae, 2011, 47(12): 92-96.
-
文章历史
- 收稿日期:2011-05-23
- 修回日期:2011-08-19
-
作者相关文章



近交系动物个体差异小、均一性好,且具有一致的基因型和遗传特征,因此是生命科学研究的试验材料。近几十年来,随着国内外线虫学研究的迅速发展,为适应不同课题研究需要,自由生活型线虫、昆虫原线虫、动物寄生及植物寄生线虫均有近交系线虫培育的报道(Dropkin et al., 1986; Gaugler et al., 1991; Sternberg et al., 2003; Bai et al., 2005; Grant et al., 2006; Dolgin et al., 2007)。
松材线虫(Bursaphelenchus xylophilus)是松树萎蔫病的病原,虽然为植物寄生线虫,却易于在实验室条件下用植物愈伤组织或真菌进行培养(Iwahori et al., 1990)。经典生物学的研究已基本阐明了松材线虫的形态结构、生活史、传播媒介、寄生范围等问题,然而,到目前为止,松材线虫病的致病机制尚未研究清楚,人们对松材线虫致病的分子机制知之甚少。分子生物学技术的发展,使得从分子水平上研究松材线虫病成为可能。已有研究表明:松材线虫群间及种群内存在高度的遗传多样性(成飞雪,2005; 陈蔚诗,2006),为此,建立遗传一致的纯合种群用于松材线虫生物学、遗传学、基因组学等研究具有重要意义。松材线虫的生殖方式为雌雄异体生殖,其繁殖可通过控制交配进行遗传操纵,基因位点近乎纯合的近交系可通过进行多次严格近交获得。本文报道了松材线虫近交系的构建过程。
1 材料与方法 1.1 供试材料试验用松材线虫为强毒虫株AMA3,该虫株由黄任娥(2007)于2004年10月分离自安徽省马鞍山市林场白马山病死松树,使用前培养于灰葡萄孢(Botrytis cinerea)上,4 ℃保存。
1.2 稀疏灰葡萄孢的培养配制1/10倍马铃薯蔗糖琼脂培养基(1/10×PDA)(马铃薯20 g,蔗糖2 g,琼脂粉15 g,水1 000 mL),于121 ℃下灭菌30 min,倒平板。将灰葡萄孢接种至上述1/10×PDA培养基中,25 ℃下培养,获得稀疏的灰葡萄孢培养基。
1.3 松材线虫AMA3虫株生活史观察首先将产卵雌虫吸入凹玻片的自来水水滴中产卵30 min,将雌虫从玻片上洗去,获得新产下的卵。添加清水至凹玻片中,于25 ℃条件下,在莱卡显微镜下每隔30 min观察卵的胚胎发育,直至孵化,获得胚胎发育时间; 进而将大量线虫置于培养皿中,产卵30 min,获得大量卵,卵孵化获得大量同步2龄线虫,将2龄幼虫接种至长满稀疏灰葡萄孢菌丝的1/10×PDA上,每小皿接30~40条,25 ℃下培养。前3天每隔12 h分离线虫,第4天每隔3 h分离线虫,在显微镜下观察线虫的发育,直至成虫出现。
1.4 松材线虫近交系的建立将松材线虫从灰葡萄孢中分离出来,用0.1%硫酸链霉素进行表面消毒(非严格无菌),用自制毛细管在显微镜下挑取性别分化明显的L4雌、雄幼虫,成对转移至用1/10×PDA培养基培养的灰葡萄孢菌块中。灰葡萄孢菌块大小为5 mm×5 mm左右,置于直径2.5~3.0 cm的小培养皿中。25 ℃下保湿培养。
根据前述生活史观察结果,于接种4~5天后,在小培养皿中加入清水,并将培养基切成更小的小块,使线虫从稀疏的灰葡萄孢中游出,在体视显微镜下检查交配情况:如果各培养皿中均未发现子代,则该近交系灭绝; 如果交配成功,但是除亲本外,出现第3条成虫,则放弃该对组合; 如果交配成功,且近交后代中无成虫出现,则从中成对挑取4龄(或3龄)线虫转移至灰葡萄孢菌块中。为了确保近交系顺利进行,每一世代尽可能多地挑取幼虫进入下一世代,剩余幼虫及亲本转入新鲜灰葡萄孢中自由繁殖,作为每一世代的母本保存。每一对亲本的后代挑完后,更换一支新的毛细管,以确保每一世代均为全同胞兄妹交配; 所有承载来自同一对亲本线虫的小培养皿放在直径15 cm的大培养皿中,以避免混乱。如此重复至20个世代,获得线虫近交系。最后,收获保留下来的近交系线虫,接种于长满灰葡萄孢的PDA培养基上,待其将葡萄孢吃尽后,再转接于玉米粒培养的灰葡萄孢中生长,保存于4 ℃冷库中。
试验过程中详细记录近交谱系。本试验中,近交系名称采用字母及数字表示,字母代表原始种群的来源及父母本代号,数字反映近交代数。具体做法是在成对挑取线虫时,给予每对线虫1个编号,然后每增加1个世代增加1个数字。
1.5 松材线虫近交系的致病力测定从冷库中将松材线虫AMA3原始种群及C、D和E近交系的第6, 10, 15和20代近交种群取出,在长满灰葡萄孢的PDA培养基上培养,10天后用贝尔曼漏斗法分离线虫,采用皮接法对2年生健康黑松(Pinus thunbergii)进行人工接种,接种量为每株2 000条,每种处理接种6株,以清水为对照。接种后24 h内给脱脂棉滴加适量蒸馏水保湿。室内温度保持25~30 ℃,逐日观察松苗发病情况。
1.6 数据分析数据采用SPSS 13.0软件进行分析。
2 结果与分析 2.1 松材线虫AMA3虫株的生活史为了能够在近交系建立过程中准确掌握每个世代分离子代幼虫的时间,首先对松材线虫AMA3虫株的各个发育阶段进行了研究,以掌握该虫株的完成1个世代所需时间,即生活史。试验结果表明:在25 ℃条件下,松材线虫从单细胞卵到2龄幼虫(L2)孵化最早为19 h,25 h后大约有60%的卵孵化。在灰葡萄孢上,2龄幼虫大约持续1天后蜕皮成为3龄幼虫(L3),3龄幼虫大约持续1天后蜕皮成为4龄幼虫(L4)。L4发育时间较长,L2接种至灰葡萄孢培养基75 h后,约有10%的成虫出现,4% L4正在蜕皮; 96 h后,成虫占23.7%,6.6% L4正在蜕皮。成虫形成后,1天之内开始交配、产卵。因此,AMA3的生活史大约为4~5天。
2.2 单对线虫交配成功率将松材线虫幼虫成对转移至灰葡萄孢块中,于25 ℃下培养,4~5天后镜检结果表明:单对线虫交配主要出现以下几种情况:1)雌、雄虫及其子代; 2)只见雌虫及其子代; 3)只见雄虫及其子代; 4)雌、雄虫均在但无子代; 5)仅见雌虫或仅见雄虫; 6)见到2条雄虫或2条雌虫。最理想的为第1种情况,在将其子代成对挑取进行近交后,雌、雄虫及剩余子代被转入新鲜灰葡萄孢中培养任其自由繁殖,作为每一世代的母本保存。第2, 3种情况可继续挑子代进行近交。如果近交成功率非常低,第4, 5, 6种情况下的雌虫和雄虫亦可重新利用,即将第4种情况下的雌虫和雄虫转移至新鲜灰葡萄孢中继续培养,第5和第6种情况下的雌虫和雄虫进行重新组合。

|
图 1 松材线虫AMA3虫株的生活史 Figure 1 Life cycle of B. xylophilus isolate AMA3 |
起初,近交繁殖是在AMA3、ZF4及GZ1 3个松材线虫原始线虫种群分别设几条线进行,但是由于工作量太大,所以放弃了ZF4及GZ1。以下仅统计起源于AMA3种群的单对线虫交配情况。
首先,从AMA3原种群中挑30对L4雌雄线虫进行单对交配,4天后,在小培养皿中加入清水,镜检结果表明,7对获得成功(成功率为23.3%)。选取其中5对子代线虫较多的开始近交繁殖,分别称A、B、C、D和E系,其中,A系和B系分别在近交第5代和第3代后灭绝。C系、D系和E系单对交配成功率分别为24.6%(536/2 182)、21.4%(257/1 201)和24.5%(406/1 657),与原始种群单对交配成功率相近。C系各世代线虫单对交配成功率差异不明显; D系和E系单对交配成功率分别于第7代和第9代出现显著下降后上升并一直保持稳定(表 1)。
|
|
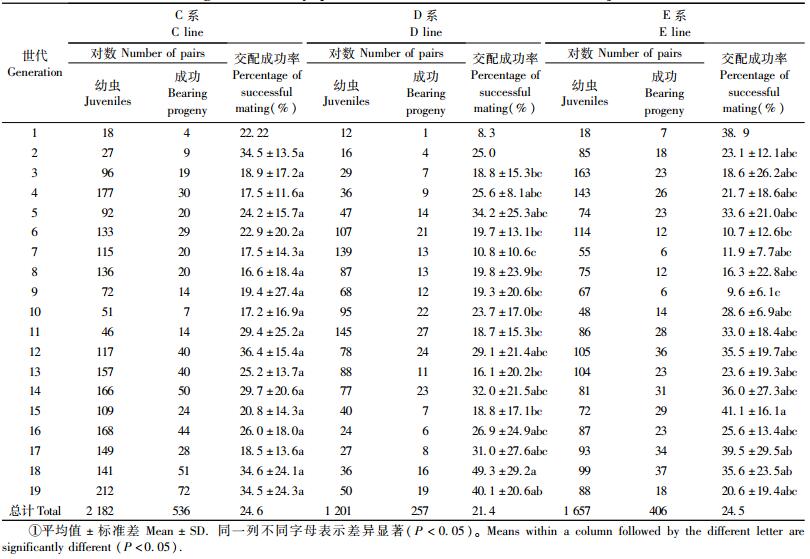
第19次近交中,共有109对线虫成功交配,产生第20代线虫(C系72对,D系19对,E系18对)(表 1),挑取了其中80对的子代至新鲜灰葡萄中扩大繁殖,最终获得近交系80个,其余交配对中因子代数量较少或已出现世代重叠而被舍弃。
2.3 松材线虫近交过程中的虫体畸形现象松材线虫近交过程中较少发现虫体畸形现象,仅在近交18代后发现6条变异虫体,其中C系和D系分别3条。虫体变异多为雌虫(5/6),主要是体态变得粗短肥胖或者体表有很多疣突,变异雌虫与正常形态雄虫交配后,其子代均恢复正常。
2.4 松材线虫近交系的致病力对2年生黑松进行接种试验,结果表明来自3对亲本的第6, 10, 15和20代近交种群线虫均能使黑松发病,有些种群的致萎率与原始种群相当,有些种群的致萎率强于原始种群其中,其中接种C系F20的植株于接种后第8天最先发病,发病率于28天后最早达100%,表明近交系线虫依然保持其强致病性。
3 结论与讨论本研究采用连续全同胞兄妹交配的方式建立了松材线虫强毒虫株AMA3的近交系。
由于线虫种类繁多,其生殖方式、生活方式各有不同。因此,近交系培育的方法也不一样。对于行孤雌生殖的线虫如花生根结线虫(Meloidogyne hapla),可通过连续每个世代转移卵块的方式进行近交系的构建(Liu et al., 2006)。雌雄同体线虫,如异小杆线虫(Heterorhabditis bacteriophora)近交系的构建可通过每世代挑取一条雌雄同体的成虫至含有共生菌的培养皿中,每一世代均确保自体交配(Gaugler et al., 1991; Bai et al., 2005)。两性生殖线虫的近交系构建相对复杂,Dolgin等(2007)通过每世代严格挑取4龄雌雄幼虫交配的方式建立了C. remanei的近交系。有些专性寄生线虫的近交系构建较为困难,如大豆孢囊线虫(Heterodera glycines)、捻转血矛线虫(Haemonchus contortus)近交系的建立(Dropkin et al., 1986; Roos et al., 2004)。
松材线虫虽为松树寄生线虫,但可实验室培养,其生殖方式为雌雄异体两性生殖,因此其近交系的构建相对简单。为了确保每一世代均为全同胞兄妹交配,本试验采取通过每世代挑取一对4龄雌雄幼虫(或3龄幼虫)交配的方式进行近交系建立,而非每世代转移一条怀孕雌虫的方式; 为了确保各近交系不出现断代,每一代均尽可能多地保持交配对数。本试验以AMA3强毒虫株为原始种群,最终建立了来自C、D和E 3对亲本的近交系,获得近交系80个。其中,C、D和E系分别挑取了2 182、1 201和1 657对线虫,其单对交配成功率分别为24.6%、21.4%和24.5%。C系各世代线虫单对交配成功率差异不明显; D系和E系单对交配成功率分别于第7代和第9代出现显著下降后上升并一直保持稳定。
通常情况下,正常异交繁殖的动植物进行近亲交配时,可致使后代受隐性、有害的特性影响,使其繁殖力、适应力降低,即近交衰退现象。Grant等(2006)发现,哺乳动物寄生线虫Parastrongyloides trichosuri近交至5~8代时,75%~80%的近交系灭绝。Dolgin等(2007)的研究表明,近交衰退使雌雄异体的C. remanei 90%的近交系灭绝,但对雌雄同体的C. elegans则出现远交衰退。本试验中,第7代和第9代出现交配成功率显著下降是否为近交衰退,不得而知。以感病黑松为宿主对AMA3原始种群及不同近交种群的致病力进行的测定结果表明,来自3对亲本的F6、F10、F15及F20松材线虫种群依然保持较强致病力,甚至有些种群的致萎率强于原始种群。当然,由于接种的黑松数量较少,只有6株,各近交种群的致病力有待进一步测定。
据笔者所查阅的文献,目前仅日本京都大学Futai教授所领导的研究小组对松材线虫进行了近交系培育,他们将强毒虫株S10和无毒虫株OKD-1近交至第10代,并以此为材料,对松材线虫虫株间致病力的遗传多样性进行了分析(Ichimura et al., 2009)。
本研究首次培育了高度近交的松材线虫近交系,这些遗传背景高度一致的群体将为松材线虫全基因组的测序发挥重要作用,也将为进一步开展松材线虫遗传、发育和生理等重要理论的研究中发挥独特作用。
陈蔚诗. 2006. 中国松材线虫群体遗传变异研究[J]. 南京:南京林业大学硕士学位论文. |
成飞雪. 2005. 松材线虫遗传多样性的AFLP研究[J]. 贵阳:贵州大学硕士学位论文. |
黄任娥. 2007. 感染松材线虫病松树滑刃目线虫研究[J]. 南京:南京林业大学博士学位论文. |
Bai C, Shapiro D I, Gaugler R, et al. 2005. Stabilization of beneficial traits in Heterorhabditis bacteriophora through creation of inbred lines[J]. Biological Control, 32(2): 220-227. DOI:10.1016/j.biocontrol.2004.09.011 |
Dolgin E S, Charlesworth B, Baird S E, et al. 2007. Inbreeding and outbreeding depression in Caenorhabditis nematodes[J]. Evolution, 61(6): 1339-1352. DOI:10.1111/evo.2007.61.issue-6 |
Dropkin V H, Halbrendt J M. 1986. Inbreeding and hybridizing cyst nematodes on pruned soybeans in petri plates[J]. Journal of Nematology, 18(2): 200-203. |
Gaugler R, Campbell J F. 1991. Selection for enhanced host-finding of scarab larvae (Coleoptera: Scarabaeidae) in an entomopathogenic nematode[J]. Environmental Entomology, 20(2): 700-706. DOI:10.1093/ee/20.2.700 |
Grant W N, Stasiuk S, Newton-Howes J, et al. 2006. Parastrongyloides trichosuri, a nematode parasite of mammals that is uniquely suited to genetic analysis[J]. International Journal for Parasitology, 36(4): 453-466. DOI:10.1016/j.ijpara.2005.11.009 |
Ichimura K, Shinya R, Takemoto S, et al. 2009. Genetic diversity in pathogenicity among isolates of the pine wood nematode, Bursaphelenchus xylophilus(Abstract)[J]. IUFRO International Symposium on Pine Wilt Disease, Nanjing, China. |
Iwahori H, Futai K. 1990. Propagation and effects of the pine wood nematode on calli of various plants[J]. Japanese Journal of Nematology, 20: 25-36. |
Liu Q L, Williamson V M. 2006. Host-specific pathogenicity and genome differences between inbred strains of Meloidogyne hapla[J]. Journal of Nematology, 38(1): 158-164. |
Roos M H, Otsen M, Hoekstra R, et al. 2004. Genetic analysis of inbreeding of two strains of the parasitic nematode Haemonchus contortus[J]. International Journal for Parasitology, 34: 109-115. DOI:10.1016/j.ijpara.2003.10.002 |