 2011, Vol. 47
2011, Vol. 47文章信息
- 杨秀艳, 孙晓梅, 张守攻, 谢允慧, 韩华
- Yang Xiuyan, Sun Xiaomei, Zhang Shougong, Xie Yunhui, Han Hua
- 日本落叶松EST-SSR标记开发及二代优树遗传多样性分析
- Development of EST-SSR markers and genetic diversity analysis of the second cycle elite population in Larix kaempferi
- 林业科学, 2011, 47(11): 52-58.
- Scientia Silvae Sinicae, 2011, 47(11): 52-58.
-
文章历史
- 收稿日期:2010-11-24
- 修回日期:2011-09-23
-
作者相关文章



采用遗传分子标记分析育种群体(优树)的遗传异质性,以遗传距离评价亲本间的遗传变异,可为了解育种群体的遗传结构、制定交配方案提供重要信息。同时,利用分子标记技术可对所收集优树群体中基因型的冗余或缺失进行评估,最终为种质资源有效群体的收集、维持、管理和提高提供数据上的支持(Leite et al., 2002)。微卫星(microsatellites)又称简单重复序列(simple sequence repeats,SSRs),在真核生物基因组中的含量非常丰富,具有多态性高、提供的信息量高、为共显性标记等优越性。EST-SSR是基于EST中简单重复序列设计的新型分子标记,不仅具备基因组SSR(gSSR)标记特点,同时更容易获得基因表达的信息,且在种间的通用性更好(忻雅等,2006; Chagné et al., 2004; Aggarwal et al., 2007)。作为植物研究中重要的遗传标记,SSRs(包括EST-SSRs)可应用于遗传图谱构建、目标基因标定、指纹图谱绘制等研究。
日本落叶松(Larix kaempferi)引入我国已有100多年,目前引种区包括温带、暖温带及北亚热带亚高山区在内的广大区域,特别是在中北亚热带亚高山区(海拔1 000~2 100 m)得到迅速发展(马常耕等,2008)。从20世纪80年代后期开始在北亚热带高山区陆续营建了日本落叶松子代测定林,目前以这些子代测定林为对象开展了日本落叶松自由授粉家系子代生长、形质与材性性状遗传变异及多性状联合选择的研究(孙晓梅等,2003; 2004a; 2004b; 2005; 杨秀艳等,2010)。在此基础上进一步选育遗传变异丰富的高世代育种材料,是当前营建日本落叶松核心育种园和二代种子园急需开展的工作,也是开展多世代遗传改良的物质基础。
以往落叶松遗传分析研究多采用随机扩增多态性的分子标记,如AFLP,ISSR,RAPD等(Arcade et al., 2000; 那冬晨等,2006; 张磊等,2008),这些标记多为显性标记,很难用于跨种或跨群体的比较基因组研究。虽也有日本落叶松SSR标记开发的报道,但所得到的多态性SSR引物并不多(19个)(Isoda et al., 2006)。本文利用作者所在课题组近年来在日本落叶松和杂种落叶松研究过程中获得的及由公共GeneBank中查询获得的落叶松EST序列开发EST-SSR标记,旨在丰富可用于落叶松遗传分析的SSR分子标记,同时利用开发的EST-SSR标记及文献检索得到的gSSR标记对北亚热带高山区选出的日本落叶松二代优树群体进行遗传变异评价,以便了解二代优树群体的遗传基础,为指导二代优树的合理利用和营建高世代种子园提供参考信息。
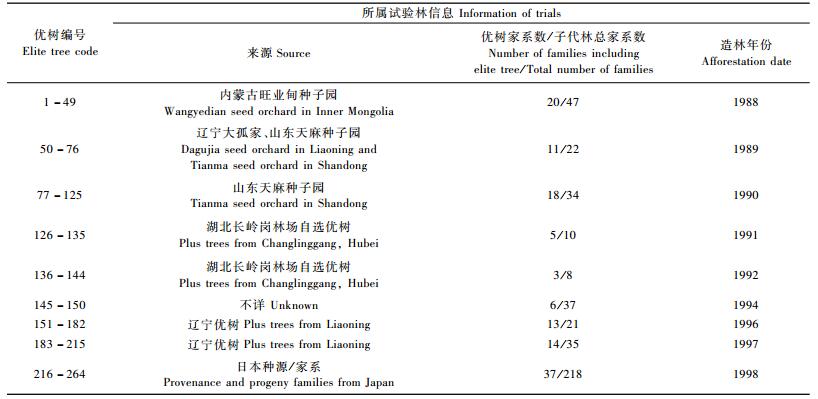
1 材料与方法 1.1 材料参试材料为2008年在湖北省建始县国有长岭岗林场日本落叶松子代测定林中依据生长指标选出的二代优树群体(杨秀艳等,2010),共计264个单株,来自121个家系,参试优树来源见表 1。2008年春季,采集264株优树完全展开的新叶置冰盒中带回实验室放于超低温冰箱(-80 ℃)中保存备用。
|
|
采用改进的CTAB法(Doyle et al., 1990)提取基因组DNA用于后续PCR扩增。
1.3 EST序列来源共计获得EST序列1 620条,来源如下:1)自NCBI公共数据库(http//:www.ncbi.nlm.nil.gov/sites/entrez)GenBank数据库中下载EST序列62条,其中源于日本落叶松木质部的有34条,登录号为AB251461-AB251494;源于日本落叶松体细胞胚组织的有28条,登录号为DN927721-DN927748。2)利用日本落叶松×华北落叶松(L. principis-rupprechtii)杂种体细胞胚(0~42天)组织为材料构建的差异表达基因文库(SSH),经测序、拼接,共获得868个UniESTs(张蕾,2008)。3)利用日本落叶松×长白落叶松(L. olgensis)2个扦插生根性状差异显著的全同胞无性系,取扦插后不同时期茎段基部材料构建的差异表达基因文库(SSH),经测序、拼接,共获得279个UniESTs(冯健,2010)。4)利用落叶松无性系1年生新梢为材料构建的茎cDNA文库,经测序、拼接,共获得411个UniESTs(李春秀,2008)。
1.4 引物设计与扩增利用SSRIT(http://www.gramene.org/db/markers/ssrtool)对1 620条EST进行SSR检索。搜索标准为:重复次数n≥4,碱基数总长度不少于12 bp,重复单元为2,3,4,5,6的核苷酸基序(motif)。PCR引物设计采用PRIMER5.0。引物设计遵循以下原则:引物长度18~24 bp; 引物退火温度50~65 ℃,最适为55 ℃,上下游引物之间退火温度不超过5℃; 扩增片段长度在50~300 bp之间; (G+C)含量为40%~65%,最适为50%。
全部SSR引物均由上海捷瑞生物工程公司合成; Taq酶、dNTPs购自泽生科技有限公司。PCR反应体系为20 μL:10×Buffer(含Mg2+)2 μL,引物浓度0.4 μmol·L-1,dNTPs浓度0.25 mmol·L-1,Taq酶浓度1 U,模板DNA30~40 ng,最后加去离子无菌水(ddH2O)至20 μL。PCR反应程序为:94 ℃预变性3 min; 94 ℃变性30 s,退火45 s,72 ℃延伸1 min,共40个循环; 72 ℃延伸7 min。
PCR产物首先采用2%琼脂糖凝胶电泳进行初步检测,舍弃无扩增产物的引物,之后在6%变性聚丙烯酰胺凝胶上电泳分离检测。将产物与6×变性上样缓冲液等体积混合,95 ℃预变性5 min后,迅速置于冰浴,取6 μL点样,在DYCZ-20C型电泳槽中进行电泳。电极缓冲液为1×TBE,65 W预电泳30 min,使凝胶板的温度达到55 ℃左右,然后65 W恒功率(君意电泳仪)电泳115 min后固定染色; 固定30 min(固定液10%乙酸),ddH2O漂洗2次,每次5 min; 用0.1%AgNO3(1 g AgNO3,1.5 mL甲醛,1 000 mL ddH2O),染色30 min,ddH2O漂洗不超过5 s,最后在预冷的显影液(30 g Na2CO3,1.5 mL甲醛,200 μL硫代硫酸钠,1 000 mL ddH2O)中显影至条带清晰,在固定液中中止显影,冲洗、干燥后扫描记录。
1.5 数据分析对目标片段进行统计,建立原始数据矩阵。利用POPGEN1.32软件包计算群体的等位基因数(A)、观察杂合度(Ho)和期望杂合度(He)。采用Nei's遗传距离(D)(Nei et al., 1979)计算样品间的遗传相似系数,采用NTSYS-pc(Rholf,2000)软件分析264个单株的遗传关系,选用SM相似系数、UPGMA法进行聚类分析。
2 结果与分析 2.1 EST-SSRs频率、类型分析在1 620条UniESTs序列中共检测到58条序列含67个SSR位点,发生频率为3.58%。在含有SSR位点的EST序列中,有4条含2个SSR重复序列,有1条含3个SSR重复序列,其余含有1个。在所发现的全部SSR重复序列中,共有二、三、五、六核苷酸4种重复类型,其中以三核苷酸为基序的SSR出现最多,共51次,占到全部SSR的76.12%;五和六核苷酸重复比较少,各占全部SSR的5.97%;二核苷酸重复占11.94%(图 1)。在二核苷酸重复中(AG)n出现频率最高; 三核苷酸重复基序种类多样,其中(AGG)n出现次数最多,为6次,其次是(AGA)n,为4次。在全部SSR重复序列中,最长的一个重复单元为二核苷酸重复单元(TA)10(AG)19。

|
图 1 落叶松EST-SSR类型及分布 Figure 1 Distribution and repeat type of EST-SSRs in larch |
全部SSR分为2种类型:一种是精确重复序列,即在串联的重复基元内无间断,数量为61个; 另一种为复合型重复序列,即2个及以上的串联基元之间有间断,且相隔的串联基元的序列组成各不相同,数量为6个。
2.2 EST-SSR引物筛选及通用性检测利用所获得的EST-SSR序列共设计引物45对,以来自不同家系的6个日本落叶松单株DNA为模板,对全部引物进行筛选。其中有14对引物没有扩增产物; 在有产物的31对引物中,有3对引物扩增片段与预期产物片段大小不符,其余28个引物中有7对引物的扩增产物具有多态性分离。表 2中列出了这7个多态位点的相关信息。
|
|

将具有多态性的EST-SSR标记在朝鲜落叶松(L. olgensis var. koreana)、长白落叶松、兴安落叶松(L. gmelinii)和华北落叶松上进行扩增,除了引物Y6和Y14在兴安落叶松上无清晰条带外,其余引物均有预期大小的条带,且具有多态性(图 2),表明所开发的EST-SSR标记在我国主要落叶松造林树种上具有良好的通用性。

|
图 2 EST-SSR引物在不同落叶松种上的扩增结果 Figure 2 Pattern of polymorphism detected by EST-SSR primer Y19, Y27 and Y85 in different larch species 1.朝鲜落叶松L. olgensis var. koreana; 2.兴安落叶松L. gmelinii; 3.长白落叶松L. olgensis; 4.华北落叶松L. principis-rupprechtii. |
试验中由于Y6仅在部分日本落叶松个体中扩增出条带,因此最终利用除Y6以外的6个EST-SSR标记和来自日本落叶松的4个SSR标记(表 3)对选出的264株日本落叶松二代优树群体进行遗传多样性分析。10对引物共扩增出46个等位基因,平均每个位点的等位基因数为4.6个。其中EST-SSR位点的平均等位基因数为3.3个,gSSR平均为6.5个,EST-SSR位点的平均等位基因数比gSSR位点明显偏少,这意味着本试验中所获得的EST-SSR位点的多态性较gSSR要低。
|
|

利用POPGEN1.32软件包从不同基因座进行等位基因观察数、观察杂合度、期望杂合度和总群体基因多样度分析(表 4)。10对引物扩出的等位基因数2~8个不等(图 3),有效等位基因平均数3.1个。平均观察和期望杂合度分别为0.590 2和0.569 1。
|
|


|
图 3 位点Y27在部分单株中的扩增结果 Figure 3 The part of amplification products of Y27 locus |
全部264个单株的遗传相似系数在0.172 5~0.966 7之间。亲缘关系最近的单株是1988年子代林中的326-1-6与1996年子代林中的98-1-3,因为这2个单株为同一家系,在遗传上有很近的亲缘关系。亲缘关系最远(遗传距离最大)的单株为1991年子代林中的365-4-5和1998年子代试验林中的149-1-5。由档案记录可知,前者为内蒙古自治区旺业甸种子园的家系子代,后者为从原产地日本引进的(草津)原生种源家系子代。依据遗传相似系数对入选二代优树群体进行聚类分析,在阈值约0.71处可将264个单株分成8个亚群体(图略),这些亚群体内个体数量相差较大,且无规律可循(如家系、种源来源相近与否等)。其中第Ⅰ亚群体内包含个体数超过全部单株数的3/5,相比之下其他亚群体内的个体较少,这表明该群体内多数单株在所检测的位点上遗传关系较近,二代优树群体内的个体数量仍有缩减空间; 另一方面,仅利用上述10个标记来检测日本落叶松二代优树群体所检测的位点数有限(46个),其代表性有一定局限性。因此,这一聚类结果只能作为亚系划分的参考依据。
3 结论与讨论 3.1 结论本文利用落叶松EST序列开发EST-SSR标记,共获得7个具有多态性和通用性标记,并在日本落叶松二代优树群体遗传多样性评价中应用。利用10个SSR标记(包括6个EST-SSR标记和4个gSSR标记)对二代优树群体264个单株的遗传差异分析表明:该群体的平均等位基因数为4.6个,有效等位基因平均数为3.1个; 平均观察杂合度和期望杂合度分别为0.590 2和0.570 2; Nei's基因多样度和Shannon多样性指数分别为0.569 1和1.096 6。这表明该优树群体具有较高的遗传变异,将其作为第2代育种群体在维持遗传多样性方面的风险较小。
3.2 讨论 3.2.1 落叶松EST-SSR标记特点及应用前景在对不同植物的研究中,EST中含有SSR重复序列的比例及类型有很大变化,如在大麦(Hordeum vulgare)、玉米(Zea mays)等谷类作物7%~10%的ESTs中含有SSR序列,其中出现最多的是三核苷酸重复,其次为二核苷酸重复(Varshoey et al., 2002),在陆地棉(Gossypium hirsutum)上发现1.1%~4.8%的EST含有SSR重复序列(Saha et al., 2003)。本研究EST序列中含有重复单元序列的比例为3.58%,略高于火炬松(Pinus taeda)和海岸松(Pinus pinaster)中1.2%和2.1%的富集比例(Chagué et al., 2004),但低于杨树(Populus) 14.8%(张新叶等,2009)的比例。这一方面是由于物种特异性引起,另一方面也可能是所采用的SSR搜索条件严格与否造成的。
由于密码子以三核苷酸为一个功能单位,因此三核苷酸位移对一个表达基因的阅读框不会造成太大影响(Metzgar et al., 2000),所以在EST序列中发现三核苷酸重复(AGG)n的SSR类型应该是最多的,本研究结果进一步印证了这一点。但有些树种的研究中出现频率最高的却是二核苷酸重复, 如杨树(张新叶等,2009)和鹅掌楸(Liriodendron)(胥猛等,2008)。这种差异可能与重复基元在ESTs中所处区域有关(ORF或UTR区域),这还有待进一步印证。本研究还发现EST-SSR标记的多态性比gSSR标记要低,以往也有类似报道(Eujayl et al., 2002)。这可能是由于EST-SSR来自功能基因的转录本,而这类序列一般相对保守,因此SSR的变异也会相应降低; 另外还可能与本研究所用的重复序列普遍较短(多为12~15 bp)有关,Cho等(2000)在水稻(Oryza sativa)的研究中发现小于18 bp的核苷酸SSR的多态性显著降低。
本研究开发的EST-SSR标记与以往研究中在落叶松上开发的gSSR标记相比,在数量和类型分布以及通用性上都具有明显的优势,可用于杂种鉴定、谱系重建、遗传作图等方面。特别需要指出的是EST-SSR属共显性标记且通用性好,与基因功能有直接关系,因此可用于比较遗传作图,以及为功能基因定位和克隆提供可行途径。
3.2.2 日本落叶松二代优树群体的遗传多样性利用10个SSR标记对日本落叶松二代优树群体进行检测发现,总群体基因多样度(H)和Shannon多样性指数分别为0.569 1和1.096 6,具有较高的遗传多样性,说明经过这一世代的选择并未造成遗传多样性的严重下降。这一结果与张薇等(2008)利用SSR标记对马尾松(Pinus massoniana)一代种子园亲本及其子代的遗传多样性水平的分析结果非常相似(种子园亲本及其子代的基因多样度分别为0.569 9和0.574 0),由此可以认为该二代优树群体基本可以代表日本落叶松第1代种子园子代的遗传多样性水平。另外,日本落叶松在引入我国后经历了百余年在不同环境下的栽培,日本落叶松与其他乡土落叶松种间、不同种源之间、个体之间易于发生天然杂交,这种基因交流也可增加其遗传多样性。
3.2.3 分子标记辅助育种和育种群体组织管理利用SSR分子标记揭示根据生长性状入选二代优树单株之间的亲缘关系可为育种群体和生产群体的组织管理提供参考。划分亚系时除根据家系信息外,还可参考单株间的亲缘关系,同一亚系内个体间的亲缘关系可以较近,但亚系间应无亲缘关系或较远,以防止和减缓近交的发生。本研究中,这批日本落叶松二代优树单株之间遗传相似系数变幅很大,说明这一群体有较宽的遗传基础。从育种群体经营的角度考虑,群体越大,经营成本就越高。本文基于SSR标记分析将264株优树划分为8个亚群体,但亚群体内个体来源并无明显规律可循。基于目前这一结果,将对每一单株进行分子标记谱带比对检测,对于谱带完全一致的单株将结合生长及材性指标等表型资料进行取舍,尽量减少亚群体内遗传相似度高的单株数,从而达到在不损失总体遗传多样性的前提下对育种群体经济、高效的经营状态。在种子园(生产群体)的配置上,亲缘关系可为一个组群的无性系配置提供参考。由于本研究所开发的落叶松SSR标记还比较少,因此对于日本落叶松个体间亲缘关系的揭示还不是十分清晰。今后将开发更多实用的SSR标记,并与AFLP或ISSR等其他标记,以及生长、材性等表型性状相结合,对日本落叶松的种质资源开展深入评价,对育种群体中个体间亲缘关系进行更细致的研究,以便为种质资源及育种群体的管理提供更为详实的依据。
冯健, 齐力旺, 孙晓梅, 等. 2010. 落叶松扦插生根过程SSH文库构建及部分基因的表达分析[J]. 林业科学, 46(6): 27-33. DOI:10.11707/j.1001-7488.20100605 |
李春秀. 2008. 日本落叶松侧枝新梢不同部位基因表达谱的比较研究[J]. 北京:中国林业科学研究院博士学位论文. |
马常耕, 孙晓梅. 2008. 我国落叶松遗传改良现状及发展方向[J]. 世界林业研究, 21(3): 58-63. |
那冬晨, 杨传平, 姜静, 等. 2006. 利用ISSR标记分析兴安落叶松种源的遗传多样性[J]. 林业科技, 31(1): 1-4. |
孙晓梅, 张守攻, 李时元, 等. 2005. 日本落叶松纸浆材优良家系多性状联合选择[J]. 林业科学, 41(4): 48-54. DOI:10.11707/j.1001-7488.20050409 |
孙晓梅, 张守攻, 侯义梅, 等. 2004a. 短轮伐期日本落叶松家系生长性状遗传参数的变化[J]. 林业科学, 40(6): 68-74. |
孙晓梅, 张守攻, 王卫东, 等. 2004b. 日本落叶松自由授粉家系形质性状遗传变异的研究[J]. 北京林业大学学报, 26(3): 41-45. |
孙晓梅, 张守攻, 齐力旺, 等. 2003. 日本落叶松自由授粉家系纸浆材材性遗传变异的研究[J]. 林业科学研究, 16(5): 515-522. |
忻雅, 崔海瑞, 卢美贞, 等. 2006. 白菜EST-SSR信息分析与标记的建立[J]. 园艺学报, 33(3): 549-554. |
胥猛, 李火根. 2008. 鹅掌楸EST-SSR引物开发及通用性分析[J]. 分子植物育种, 6(3): 615-618. |
杨秀艳, 张守攻, 孙晓梅, 等. 2010. 北亚热带高山区日本落叶松自由授粉家系遗传测定与二代优树选择[J]. 林业科学, 46(8): 45-50. DOI:10.11707/j.1001-7488.20100807 |
张磊, 张含国, 李雪峰, 等. 2008. 落叶松种间及无性系间ISSR鉴别技术的研究[J]. 植物研究, 28(2): 216-221. |
张蕾. 2008. 日本落叶松×华北落叶松体细胞胚胎发生的生化机制和分子机理研究[J]. 北京:中国林业科学研究院博士学位论文. |
张薇, 龚佳, 季孔庶. 2008. 马尾松实生种子园遗传多样性分析[J]. 分子植物育种, 6(4): 717-723. |
张新叶, 宋丛文, 张亚东, 等. 2009. 杨树EST-SSR标记的开发[J]. 林业科学, 45(9): 53-59. DOI:10.11707/j.1001-7488.20090910 |
Arcade A, Anselin F, Rampant P F, et al. 2000. Application of AFLP, RAPD and ISSR markers to genetic mapping of European and Japanese larch[J]. Theor Appl Genet, 100(2): 299-307. DOI:10.1007/s001220050039 |
Aggarwal R K, Hendre P S, Varshney R K, et al. 2007. Identification, characterization and utilization of EST-derived genic mierosatellite markers for genome analyses of coffee and related species[J]. Theor Appl Genet, 114(2): 359-372. DOI:10.1007/s00122-006-0440-x |
Cho Y G, Ishii T, Temnykh S, et al. 2000. Diversity of microsatellites derived from genomic libraries and GenBank sequences in rice (Oryza sativa L[J]. ). Theor Appl Genet, 100(5): 713-722. DOI:10.1007/s001220051343 |
Chagné D, Chaumeil P, Ramboer A, et al. 2004. Cross-species transferability and mapping of genomic and cDNA SSRs in pines[J]. Theor Appl Genet, 109(6): 1204-1214. DOI:10.1007/s00122-004-1683-z |
Doyle J J, Doyle J L. 1990. Isolation of plant DNA from fresh tissue[J]. Focus, 12: 13-15. |
Eujayl I, Sorrels M E, Baum M, et al. 2002. Isolation of EST-derived microsatellite markers for genotyping the A and B genomes of wheat[J]. Theor Appl Genet, 104(2/3): 399-407. |
Isoda K, Watanabe A. 2006. Isolation and characterization of microsatellite loci from Larix keampferi[J]. Molecular Ecology Notes, 6(3): 664-666. DOI:10.1111/men.2006.6.issue-3 |
Leite S M M, Bonine C A, Mori E S, et al. 2002. Genetic variability in a breeding population of Eucalyptus urophylla S[J]. T. Blake. Sivae Genetica, 51(5/6): 253-256. |
Metzgar D, Bytof J, Wills C. 2000. Selection against frameshift mutation limits microsatellite expansion in coding DNA[J]. Genome Res, 10(1): 72-80. |
Nei M, Li W H. 1979. Mathematical model for studying genetic variation in term of restriction endonuclease[J]. Proc Natl Acad Sci USA, 76(10): 5269-5273. DOI:10.1073/pnas.76.10.5269 |
Rohlf J F. 2000. NTSYS pc: Numerical Taxonomy and Mulitivariate Analysis System[J]. Version 2.1, Users Guide. Exeter Software, Setauket, New York. |
Saha S, Karaca M, Jenkins J N, et al. 2003. Simple sequence repeats as useful resources to study transcribed genes of cotton[J]. Euphytica, 130(3): 355-364. DOI:10.1023/A:1023077209170 |
Varshoey R K, Thiel T, Stein N, et al. 2002. In silico analysis on frequency and distribution of microsatellites in ESTs of some cereal species[J]. Cell Mol Biol Lett, 7(2A): 537-546. |