 2011, Vol. 47
2011, Vol. 47文章信息
- 徐红霞, 辛中尧, 王洪建, 王香枝
- Xu Hongxia, Xin Zhongyao, Wang Hongjian, Wang Xiangzhi
- 甘肃白水江自然保护区的天牛群落多样性
- Community Diversity of Longhorn Beetles in Baishuijiang Nature Reserve
- 林业科学, 2011, 47(8): 182-187.
- Scientia Silvae Sinicae, 2011, 47(8): 182-187.
-
文章历史
- 收稿日期:2009-12-18
- 修回日期:2010-04-02
-
作者相关文章


生物多样性的研究和保护是维持生态平衡的关键,昆虫多样性又是生物多样性的重要组成部分(Samways, 1993;王献溥等,1994),有的学者甚至认为昆虫主宰着全球的生物多样性(Lawton, 2002)。国内外已有大量有关昆虫群落多样性的研究,而且大多是结合害虫管理开展的,其内容包括群落的组成结构特征、各级营养水平分析以及植物群落结构变化对昆虫群落结构和生态效应的影响等(戈峰,2008;Haddad et al., 2001;郝树广等,2000)。天牛是一类重要的蛀干害虫,全世界已知4 500种以上,我国已知2 000种以上,几乎所有的针叶树和阔叶树都不同程度地受到天牛的为害(张执中,1977)。研究表明:天牛具有应用于生物多样性评价方面的潜力(Allison et al., 2004)。甘肃白水江国家级自然保护区是世界最大的野生大熊猫保护区,区内植被类型多样,天牛种类十分丰富,笔者于2007—2009年对白水江自然保护区境内天牛的种类、多样性变化进行了系统调查和研究,以期为评价保护区生物多样性提供一些基础材料。
1 研究区概况甘肃白水江国家级自然保护区位于甘肃省最南端,我国亚热带北缘,是甘肃省物种最丰富的地区,地理位置在104°16'—105°27'E,32°16'—33°15'N之间,主体部分为白龙江以南的岷山山系摩天岭北坡区域以及摩天岭南坡碧口李子坝区域。海拔595 ~ 4 072 m, 年均温15.6 ℃,雨量充沛,年降水量500 ~ 1 000 mm, 无霜期270天左右。由于受地形的影响,当河谷地区为盛夏季节时,高山仍有白雪覆盖,形成了该地区独特的植被结构。植被的种群分布,从水平方面看,东南碧口中庙等地种类较多,西部种类较少,常绿阔叶林仅分布在东端,且具有珍稀濒危种类多的特点。从垂直方面来看,有着完整的山地植被垂直带(王洪建等,1997)。
2 研究方法 2.1 调查方法根据自然保护区内的地貌及植被类型,参考相关文献(王洪建等,1997),按海拔595 ~1 100 m, 1 100 ~ 1 600 m, 1 600 ~ 2 100 m, 2 100 ~2 900 m, 2 900 ~ 3 450 m, 3 450 m以上等6个海拔区间将生境划分为常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、针阔混交林、亚高山针叶林、高山灌丛草甸6种类型。在保护区内按海拔和植被类型设置样带,每种植被类型设置2个样带,共12个样带,每个样带长不小于5 km。
2007—2009年,每年6—8月天牛成虫羽化期,沿样带步行,行走速度约为1 ~ 1.5 km·h-1,以目光搜索和扫网相结合的方法进行采集,并在每个样带中选1个有代表性的样点,夜间架设250 W汞灯,选用2.5 m × 2.0 m白布作为幕布,清除幕布四周4 ~5 m内的杂草,晚上9: 00—12: 00时开灯诱集具有趋光性的成虫。对容易识别、常见的种类,采用直接观察法统计种类和数量,不能识别种类的成虫用乙酸乙酯毒杀,编号后,置于储虫瓶中,带回室内鉴定分析。
2.2 种类鉴定主要依据嘉理思1)、华立中(2009)、蒋书楠等(2001)的分类系统以及近年来我国出版的天牛科分类专著进行分类(周嘉熹等,1988)。
1) 嘉理思. 1983.中国天牛科检索表.华立中,译.中山大学印.
2.3 数据处理各类群数量优势度的划分:以特定类群个体数占所有类群总个体数的百分比(D)作为划分标准,D≥10%为优势类群,1% ≤D<10%为常见类群,D<1%为稀有类群;优势种优势度采用Berger-Parker指数(Dn)表示,按以下计算公式:Dn= Nmax/ NT,其中,Nmax为优势种的种群数量,NT为群落全部种类的种群数量(戈峰,2008)。反映群落的多样性指数有很多,但是每个多样性指数反映多样性不同方面(Shimatani et al., 2004),本研究中物种丰富度直接用物种数(S)来表示,采用ShannonWiener指数(H)和Simpson指数(D)、Pielou均匀度指数(J)分析天牛群落的特征,用相异系数(1 -Cs)比较不同植被类型中天牛群落的相异程度(孙儒泳,2000;王敏等,2003),计算公式如下:
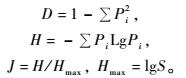
|
式中: Pi为第i种的个体数与总个体数的比值,S为物种数。
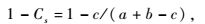
|
式中: c为2个群落或样地共有的物种数,a和b分别为样地A和样地B的物种数。
利用Excel, DPS统计软件进行以上统计分析。
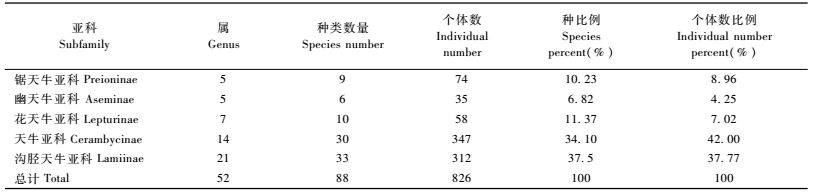
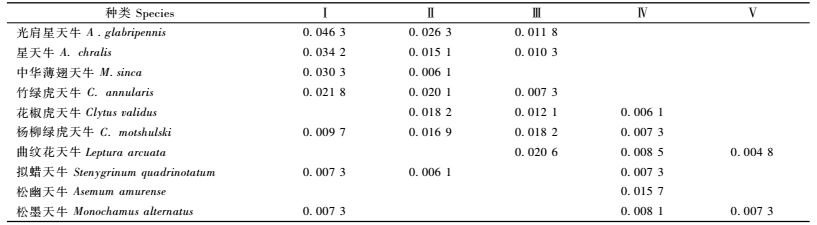
3 结果与分析 3.1 天牛群落的总体特征调查过程中共采集天牛826只,隶属5个亚科88种(表 1及附表),天牛亚科和沟胫天牛亚科为优势亚科,2个亚科分别占天牛总个体数的42%和37.77%,合计占天牛总个数的79.77%;锯天牛亚科、花天牛亚科和幽天牛亚科为常见类群,合计占总个数的20.23%;星天牛属,绿虎天牛属,白条天牛属和颈天牛属为优势属,合计占总个数的38.98%;优势种(表 2)的优势并不明显,数量最多的种是光肩星天牛(Anoplophora glabripennis),优势度为0.080,其次依次是星天牛(A.chralis)、杨柳绿虎天牛(Chlorophorus motshulski)、云斑白条天牛(Batocera horsfieldi)等。
|
|
|
|

1) 群落组成多样性 随着海拔和植被类型的不断变化,天牛群落也发生相应的变化。从表 3可以看出:天牛群落中亚科、种的丰富度、多样性指数(H)和优势度指数(J)均随海拔的增加和植被类型由常绿阔叶林带、落叶阔叶林到针叶林带的变化,呈明显减少的趋势,低海拔595 ~ 1 100 m的常绿阔叶林带种的丰富度、多样性指数(H)和优势度指数(J)均为最大,分别为69,5.667 5和0.975 9;而高海拔2 900 ~ 3 450 m的寒温性针叶林带多样性指数和优势度指数均为最小,分别为15,0.704 8和1.565 6。在海拔900 ~ 1 600 m, 植被类型为常绿阔叶林和常绿落叶阔叶混交林的范围内,天牛科5个亚科均有分布,这2个林带分别采集到69种、440个和35种、234个天牛,占采集天牛总数的81.6%,表明大多数天牛类群分布在海拔比较低的常绿阔叶林带和常绿落叶阔叶混交林带;在海拔2 100 ~3 450 m的针阔叶混交林带和寒温性针叶林带中,由于海拔高,植物种类减少,天牛种类和数量急剧减少,只采集到10种、51个天牛,多样性指数也最小;在海拔3 450 m以上的高山灌丛、高山草甸带则没有采集到天牛。
|
|
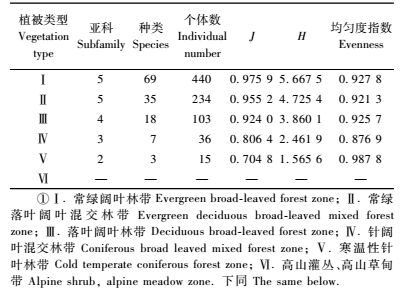
不同植被类型中的天牛群落均匀度也不一样,从表 3看出:海拔2 900 ~ 3 450 m的寒温性针叶林带天牛群落均匀度最高,其次是海拔分别为595 ~1 100 m的常绿阔叶林带和1 600 ~ 2 100 m的落叶阔叶林带,均匀度最低的是植被类型由常绿阔叶林向落叶阔叶林过渡的海拔1 100 ~ 1 600 m的常绿、落叶阔叶混交林带和植被类型由落叶阔叶林向针叶林过渡的海拔2 100 ~ 2 900 m的针阔叶混交林带。
2) 优势种及其优势度 从表 4可以看出:中华薄翅天牛、星天牛、光肩星天牛是常绿阔叶林带的优势种,中华薄翅天牛只分布在低海拔的植被类型中,海拔1 600 m以上,就很少见到分布;星天牛、光肩星天牛分布较广,在海拔2 100 m以下的植被类型中均有分布;竹绿虎天牛、花椒虎天牛和杨柳绿虎天牛是常绿、落叶阔叶混交林带优势种,分布比较广泛,尤其是杨柳绿虎天牛,在海拔2 900 m以下的各植被类型中均有分布,同时也是落叶阔叶林带优势种之一;落叶阔叶林带的其他优势种还有曲纹花天牛;松幽天牛是阔叶混交林带的优势种;松墨天牛为寒温性针叶林带的优势种,适应性也很广,从低海拔595 ~ 1 100 m到高海拔2 900 ~ 3 450 m的植被类型中均有分布。
|
|
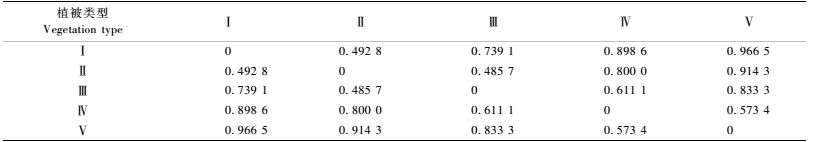
3) 天牛群落的相异性 通过计算5种不同海拔的植被类型中天牛群落的相异性系数(表 5),并采用相异性系数最短距离法对5种植被类型的天牛群落进行了聚类分析(图 1)。结果表明:不同植被类型的天牛群落间相异性较高,常绿、落叶阔叶混交林带和落叶阔叶林带天牛群落相异性最低,在相异系数0.485 7时聚为一类;其次是常绿阔叶林与常绿、落叶阔叶混交林带的天牛群落,在相异系数0.492 8时聚为一类;针阔叶混交林带和寒温性针叶林带天牛群落在相异系数0.571 4时聚为一类;最后与前3个群落在相异系数0.611 1时聚为一类,说明从常绿阔叶林带向落叶阔叶林带过渡的常绿、落叶阔叶混交林带的天牛群落与落叶阔叶带天牛群落相异性较低,针阔叶混交林带的天牛群落多样性与寒温性针叶林带的天牛群落之间相异性低,表明它们之间更为相似。
|
|

|
图 1 不同植被类型的天牛群落聚类分析 Figure 1 Cluster analyses of longhorn beetles communities in different vegetation types |
在甘肃白水江国家级自然保护区内共采集到天牛88种,分隶于5亚科52属,反映了该地区在科、属、种各级水平上的多样性均较高,天牛科的种类占目前甘肃省记录(166种)1)的53.01%,这充分体现了白水江自然保护区的生物多样性的特点。
1) 张友,邴积才. 1999.甘肃省第2次森林病虫普查名录.甘肃省森林病虫防治检疫站编,186 -506.
天牛是植食性昆虫,幼虫营钻蛀性生活,主要以木本植物为寄主,且寄主范围比较窄,因此植被类型对天牛群落的组成和数量变化有直接的影响(黄建华等,2003)。甘肃白水江国家级自然保护区主体部分地处岷山支脉摩天岭阴坡或半阴坡,有完整的山地植被垂直带(王洪建等,1997)。植被的垂直分布也表现在天牛群落多样性的变化中,天牛群落多样性与植被垂直带相关,随着海拔的增加,植物群落结构的简单化,天牛种类、群落多样性明显减少。在高海拔的针阔叶混交林带、寒温性针叶林带以及高山灌丛、高山草甸采集到的天牛种类和数量很少,甚至没有采集到天牛成虫,这可能是植被单一,天牛种类和数量原本就少,也有可能是由于样带选择不尽合理,这有待于进一步研究和改进。
物种丰富度格局随海拔梯度变化是一直是生态学研究的重要议题(Whittaker et al., 1960;Zhao et al., 2005)。尤其是在山区,海拔是控制不同物种格局的主要因素。物种多样性随海拔梯度的变化格局是了解山区生态系统功能并对其进行合理管理的重要依据(井学辉等,2010)。目前山地生态系统研究中,约75%的研究显示生物多样性从低海拔到高海拔呈单峰或偏锋分布格局,在中海拔处生物多样性达到顶峰(Grytnes, 2003;刘洋等,2009)。黄建华等(2003)在广西猫儿山天牛群落调查中,中海拔600 ~ 1 200 m区间,天牛多样性也是最高。可能由于白水江自然保护区中海拔1 100 ~ 1 600 m常绿落叶阔叶混交林分多位于保护区外缘,人为干扰严重,部分已被农田替代(王洪建等,1977),致使木本植物数量降低,从而使天牛的多样性随海拔升高而减少,在中海拔区域并未表现出种类和数量上的优势。这可能也是位于海拔1 100 ~ 1 600 m常绿落叶阔叶混交林带的天牛群落与海拔1 600 ~ 2 100 m落叶阔叶混交林带天牛群落更为相似的原因。
附录Addenda:|
|

戈峰. 2008. 昆虫生态学原理与方法[M]. 北京: 高等教育出版社: 254-255.
|
郝树广, 张孝曦, 程遐年. 2000. 稻田节肢动物群落优势功能集团的垂直分布、数量动态及天敌作用估计[J]. 应用生态学报, 41(4): 343-353. |
华立中, 奈良一, 塞缪尔林G A. 2009. 中国天牛彩色图鉴[M]. 广州: 中山大学出版社.
|
黄建华, 周善义. 2003. 广西猫儿山天牛科昆虫多样性研究[J]. 广西师范大学学报:自然科学版, 21(3). |
蒋书楠, 陈力. 2001. 中国动物志. 昆虫纲. 第二十一卷, 鞘翅目: 天牛科: 花天牛亚科. 北京: 科学出版社.
|
井学辉, 臧润国, 丁易. 2010. 新疆阿尔泰山小东沟北坡植物多样性沿海拔梯度分布格局[J]. 林业科学, 46(1): 23-28. DOI:10.11707/j.1001-7488.20100104 |
刘洋, 张健, 杨万勤. 2009. 高山生物多样性多气候变化的响应的研究进展[J]. 生物多样性, 17(1): 88-96. |
孙濡泳. 2001. 动物生态学原理[M]. 3版. 北京: 北京师范大学出版社: 396.
|
王洪建, 武光和. 1997. 甘肃白水江国家级自然保护区综合科学考察报告[M]. 兰州: 甘肃科学技术出版社.
|
王敏, 黄国华, 范骁凌, 等. 2003. 石门台自然保护区蝴蝶多样性研究[J]. 生物多样性, 11(6): 441-453. |
王献溥, 刘玉凯. 1994. 生物多样性理论和实践[M]. 北京: 中国环境科学出版社.
|
张执中. 1977. 森林昆虫学[M]. 北京: 中国林业出版社: 321-322.
|
周嘉熹, 孙益知, 唐鸿庆. 1988. 陕西省经济昆虫志. 鞘翅目: 天牛科. 西安: 陕西科学技术出版社.
|
Allison J D, Borden J H, Seybold S J. 2004. A review of the chemical ecology of the Cerambycidae (Coleoptera)[J]. Chemoeeology, 14(3 /4): 123-150. |
Grytnes J A. 2003. Species-richness patterns of vascular plants along seven altitudinal in Norway[J]. Eeography, 26(3): 291-300. |
Haddad N H, Tilman D, Haarstad J, et al. 2001. Contrasting effects of plant richness and composition on insect community: a field experiment[J]. The American Naturalist, 158: 17-35. DOI:10.1086/320866 |
Lawton J H. 2002. Abstract of international congress of entomology Londrina: Embrapa Soja.
|
Samways M J. 1993. Insect in biodiversity conservation: some perspective and directives[J]. Biodiversity and Conversation, 2(3): 258-282. |
Shimatani K, Kubota Y. 2004. Quantitative assessment of multispecies spatial pattern with high species diversity[J]. Ecological Research, 19(2): 149-163. DOI:10.1111/ere.2004.19.issue-2 |
Whittaker R J, Willis K J, Field R. 1960. Vegetation of the Siskiyou Mountains, Oregon and California[J]. Ecological Monograph, 30(3): 279-338. DOI:10.2307/1943563 |
Zhao C M, Chen W L, Tian Z Q, et al. 2005. Altitudinal pattern of plants species diversity in Shennongjia Mountains, central China[J]. Journal of Integrative Plant Biology, 47(12): 1431-1449. DOI:10.1111/jipb.2005.47.issue-12 |