 2011, Vol. 47
2011, Vol. 47文章信息
- 李振华, 于澎涛, 王彦辉, 李志勇, 王轶浩, 杜阿朋
- Li Zhenhua, Yu Pengtao, Wang Yanhui, Li Zhiyong, Wang Yihao, Du Apeng
- 重庆酸雨区受害马尾松林凋落物特征及其环境因子响应
- Characters of Litter-Fall in Damaged Pinus massoniana Forests and Its Responses to Environmental Factors in the Acid Rain Region of Chongqing, China
- 林业科学, 2011, 47(8): 19-24.
- Scientia Silvae Sinicae, 2011, 47(8): 19-24.
-
文章历史
- 收稿日期:2010-05-06
- 修回日期:2011-06-20
-
作者相关文章



2. 河南科技大学农学院 洛阳 471003;
3. 国家林业局桉树研究开发中心 湛江 524022
2. College of Agriculture, Henan University of Science and Technology Luoyang 471003;
3. China Eucalypt Research Centre Zhanjiang 524022
凋落物是林木生长发育的代谢产物, 对养分归还和地力维持有重要意义, 也是林木对干旱等常见胁迫(杨帆等, 2007)的响应指标。国内外学者曾报道过低温干旱加速杉木(Cunninghamia lanceolata)针叶枯黄(温远光等, 1987)、极端干旱促进植物叶片衰老和凋落(Meentemeyer et al., 1982)。探讨凋落物特征及其对环境变化的响应机制, 是森林养分循环和森林健康研究的重要内容。
受地理地形、能源结构及社会因素等的影响, 重庆市长期遭受严重的大气污染和酸雨危害, 酸雨强度大、频率高、pH值低, 导致当地主要造林树种马尾松(Pinus massoniana)生长量下降、冠层稀疏、针叶脱落和根系严重退化(李志勇等, 2009)。一般来说, 根系减少到一定程度必然使林木对土壤水分亏缺变得敏感, 进而影响树叶凋落、林木生长和森林健康。随着全球气候变化导致的高温、干旱等灾害性极端气象事件的频繁发生, 森林生态系统的水分养分循环及健康状况必然受到极大影响, 因此非常有必要及早地开展和加强受害森林对环境变化的响应机制研究。
本研究在重庆酸雨区铁山坪林场选择典型受害马尾松林建立固定样地, 同步监测生长季的气象、土壤含水量和凋落物数量及其组分的时间变化, 分析凋落动态与气象和土壤水分条件的关系, 为深入认识酸化胁迫下森林生态系统的健康状况并制定相应的科学管理决策提供理论依据。
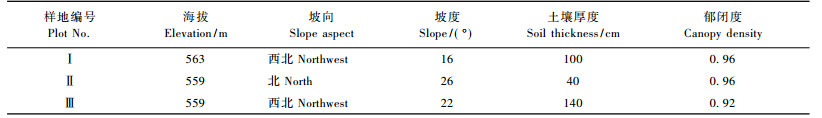
1 研究区概况研究区位于重庆市东北部的铁山坪林场(106° 41'E, 29°38'N), 海拔512~579 m, 属亚热带湿润季风气候, 冬季温暖, 夏季炎热, 年均气温18 ℃, 年均空气相对湿度80%, 年均降水量1 100 mm, 但夏季降水不均且多发伏旱(http:∥ www.121.cq.cn/ service/ cqweather.htm)。土壤为砂岩上发育的山地黄壤, 厚50~80 cm, 密度1.25~1.34 g·cm-3, 总孔隙度47%~50%, pH值3.6~4.2 (杨永森等, 2006)。主要森林类型为马尾松天然次生林, 其主林冠层几乎完全由马尾松组成, 冠下混生少量杉木、木荷(Schima superba)、樟树(Cinnamomum camphora)和毛桐(Mallotus barbatus)等; 灌木层主要植物有油茶(Camellia oleifera)和山黄皮(Randia cochinchinensis)等; 草本层以蕨类为主, 如芒萁(Dicranopteris pedata)、狗脊蕨(Woodwardia japonica)和黑足鳞毛蕨(Dryopteris fuscipes)等。由于长期遭受酸沉降和土壤酸化危害, 马尾松林的健康状况和生产力明显下降(李志勇等, 2009), 针叶脱落率40%~50%, 个别年份的林木死亡率高达6.1% (冯宗炜, 2000)。
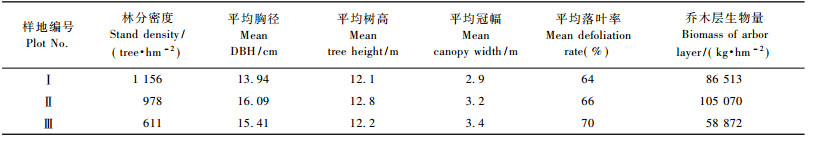
2 研究方法在林龄为50年且林相较整齐的马尾松纯林中, 设立3块30 m × 30 m的定位监测样地(表 1)。对样地内胸径(DBH) ≥ 4 cm的树木进行每木调查, 包括种类、胸径(cm)、树高(m)、枝下高(m)、冠幅(m)和树冠落叶率(标度单位为5%)等指标, 并根据马尾松全株生物量回归方程计算乔木层生物量, 回归方程为W = aDbN(a = 0.097 7, b= 2.520 6, R2 = 0.99), W为乔木层生物量(kg·hm-2), D为平均胸径(cm), N为林分密度(tree·hm-2) (张治军等, 2006), 结果见表 2。
|
|
|
|
在每个样地选取5个典型地点(避开灌木遮蔽和林窗, 冠层厚度适中)安装5个凋落物收集器。收集器由1 mm孔径的尼龙网制成, 大小为1 m × 1 m × 0.3 m, 四周固定在木桩上, 底部距地面0.8 m。2009年3月1日—11月30日, 每周周末将收集器内的凋落物样品全部收回, 同一样地的样品统一混合、风干, 分拣出松针、松枝、松花(马尾松雄花序)、树皮、松果(马尾松球果)及其他(伴生树种的枝叶、虫鸟粪便或残体)等组分, 将各组分在65 ℃下烘干至恒质量, 计算凋落量。
2009年3—11月, 每月的月初和月末分别在各样地内用土钻采取土壤样品, 每隔10 cm为一层, 每层3次重复, 取至母岩层, 然后将土样带回室内, 在105 ℃下烘干至恒质量, 并根据各层土壤密度实测数据计算各样地土壤体积含水量。利用3个样地的土壤含水量平均值作为为马尾松林分的土壤含水量, 每月2次取样的平均值为当月土壤含水量。
在研究区的林外开阔处, 安装1台WeatherHawk-232自动气象站, 连续监测降水量(mm)、蒸发量(mm)、空气温度(℃)、空气相对湿度(%)、风速(m·s-1)等气象指标, 每隔5 min自动采集1次数据; 另设1个雨量筒和1个蒸发皿(内径均为20 cm), 辅助测量降水量和蒸发量。降水量和蒸发量监测从2009年3月1日开始, 其余气象指标监测从5月1日开始, 11月30日结束。
采用Microsoft Excel和SPSS 11.5统计软件进行数据处理分析。
3 研究结果 3.1 气象因子和土壤含水量变化气象观测结果表明(表 3):研究期间共降水1 287 mm, 其中62%集中在夏季(6—8月), 25%集中在春季(3—5月), 13%集中在秋季(9—11月)。春、秋季的月降水量变动较小, 夏季变动剧烈。7月份为当地伏旱期, 高温少雨, 仅降水62 mm。生长季总蒸发量为1 249 mm, 月蒸发量在3—8月逐月递增, 之后快速下降。月均气温与相对湿度的变化趋势相反, 2者的变动范围分别为10.81~26.79 ℃和53.01%~82.61%。月均风速2.08~3.80 m·s-1, 5—9月波动上升, 9月之后逐月下降。
|
|
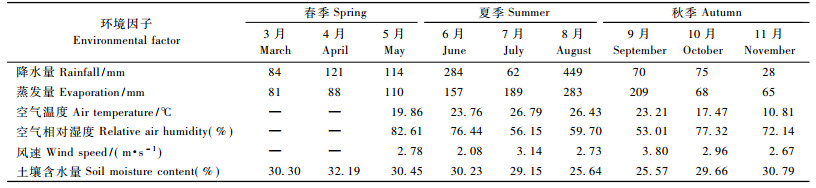
马尾松林土壤含水量在春季保持较高水平, 变动范围为30.30%~32.19%;入夏以后, 随着气温、风速和蒸散的增高, 土壤含水量逐渐由6月的30.23%降至8月的25.64%; 9月最低, 为25.57%, 当月空气湿度为生长季内最低, 而风速达到最高; 之后随着气温、蒸散等的降低, 土壤含水量回升至较高水平。6和8月的降水量很大, 但暴雨产生了大量地表径流, 降低了其对土壤水分恢复的贡献。按季节评价时, 春季土壤含水量最高(30.98%), 秋季次之(28.60%), 夏季最低(28.34%)。
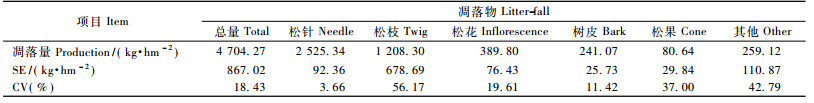
3.2 凋落物特征由表 4可见:生长季马尾松林的总凋落量为4 704.27 kg·hm-2, 变异系数为18.43%。凋落物中比例最大的组分是松针(53.68%), 其次是松枝(25.69%), 而松花(8.29%)、树皮(5.12%)、松果(1.71%)和其他(5.51%)的比例较小。样地间的变动以松针为最小, 变异系数仅为3.66%, 属弱变异; 其余组分均属中等变异。
|
|
月凋落量呈明显的季节性变化(表 5)。整体来看, 5—6月为凋落低谷期, 7—11月为高峰期。春季(3—5月)凋落较少, 占生长季总量的24.96%, 且月际、样地间的变动都很小; 入夏以后(6—8月), 月凋落量迅速升高, 并于初秋(9月)达到峰值; 秋季(9—11月)凋落最多, 占生长季总量的48.33%, 且月际、样地间的变动最大, 10—11月的样地间变异系数均超过了50%。
|
|
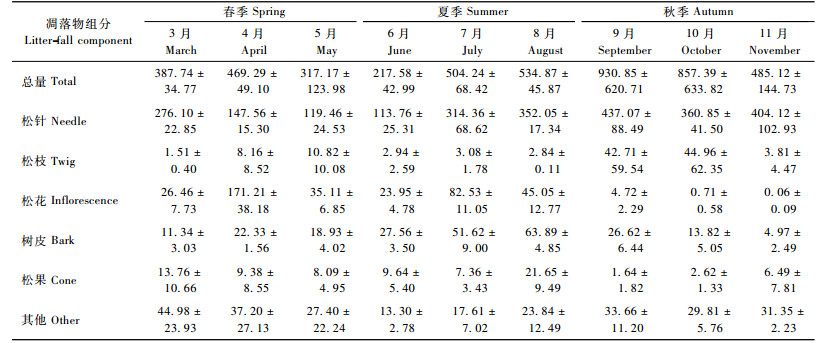
受酸雨和土壤酸化影响, 研究区内马尾松树冠上极少存留多年生针叶, 所以生长季凋落的松针多为1年生的。表 5显示:松针凋落量在3—6月逐月减少, 6月仅为113.76 kg·hm-2; 7月开始迅速增多, 并保持较高水平至11月。整体来看, 4—6月为凋落低谷期, 月均凋落量小于150 kg·hm-2; 7—11月为高峰期, 月凋落量314.36~437.07 kg·hm-2。值得注意的是, 6—9月的松针凋落量逐月上升是伴随土壤含水量持续下降而发生的。由此推断, 土壤水分相对不足造成的根系受害、细根减少且分布变浅、林木蒸腾增加可能是加剧马尾松针叶凋落的重要原因。松枝凋落的季节差异性非常明显, 春夏两季凋落量之和不足生长季总量的1/3, 秋季凋落量及月际变动均为最大, 9—10月大量松枝集中脱落, 很可能与8—9月土壤含水量持续降低导致的枝条干枯以及后期的降水、大风等外力作用有关(卢立华等, 2008)。树皮凋落的月际变化呈单峰型曲线(8月最高), 季节分配表现为春秋季少, 夏季多。松花即马尾松雄花序, 形成于早春, 凋落多在4月, 部分残留至7月形成凋落次高峰。松果凋落多在初春(3月)和夏末(8月)。凋落物中的其他组分来源较杂, 规律性不强, 但春秋季多、夏季少。
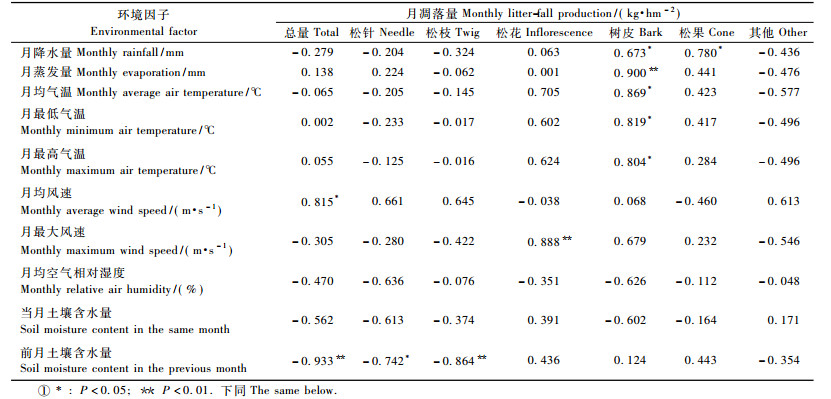
3.3 凋落量与环境因子的相关性由表 6可见:月凋落总量与前月土壤含水量极显著负相关(P<0.01), 并与月均风速显著正相关(P<0.05), 而与其他环境因子相关不显著(P>0.05)。月凋落松针量、松枝量与前月土壤含水量分别呈显著、极显著负相关。这表明凋落总量及其主要组分(松针、松枝)的凋落量明显受土壤湿度影响, 但其对土壤湿度的响应存在时间滞后。月凋落松花量与月最大风速显著正相关。月凋落树皮量与月蒸发量极显著正相关, 与月均气温、月最低和月最高气温、月降水量均显著正相关, 表明树皮脱落很可能是由高温干旱使树皮翘裂和降水使树皮增重共同导致的。月凋落松果量与月降水量显著正相关。月凋落其他量与各环境因子相关不显著。
|
|
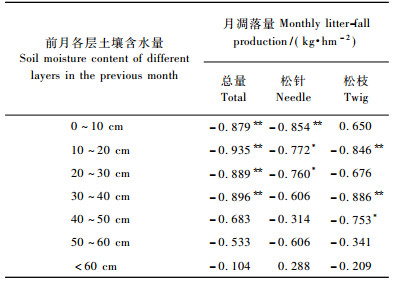
另经相关分析得知:7个月凋落量指标与当月各土层含水量均不相关(相关系数表略), 且与前月各土层含水量的相关关系各有不同(表 7)。月凋落总量与前月0~40 cm各土层含水量极显著负相关, 与40 cm以下各土层相关不显著。月凋落松针量受前月0~30 cm各土层含水量的显著影响, 尤其与0~10 cm土层极显著负相关, 表明松针凋落对土壤干旱非常敏感。月凋落松枝量受中上土层含水量的显著影响, 尤其与30~40 cm和10~20 cm土层含水量极显著负相关。其余各组分月凋落量指标与前月各土层含水量相关均不显著(表略)。
|
|
为探讨月凋落量随前月特定土层含水量变化的趋势, 选取月凋落总量、松针量和松枝量与其相关最紧密的土层含水量分别建立回归曲线(图 1)。可见, 月凋落松针量随0~10 cm土层含水量降低而近线性升高, 并在含水量降至28%时趋于平稳; 月凋落松枝量则在30~40 cm土层含水量低于28%后急剧升高; 月凋落总量随10~20 cm土层的含水量降低而近线性升高。

|
图 1 月凋落量与前月特定土层含水量的关系 Figure 1 Relationship between monthly litter-fall production and soil moisture content of certain layer in the previous month |
重庆酸雨区受害马尾松林的生长季(3—11月)凋落物总量为4 704.27 kg·hm-2, 主要由松针和松枝组成, 2者分别占总量的53.68%和25.69%, 松花、树皮、松果和其他组分的比例较小。主要受表层土壤湿度和风速的影响, 马尾松林在5—6月为凋落低谷期, 7月进入高峰期, 并持续到生长季结束。凋落物总量表现为秋季>夏季>春季。松针凋落高峰为7—11月, 松枝凋落集中在9—10月, 2者仅受表层土壤水分的显著影响; 受生理节律和月最大风速影响, 松花多在4和7月凋落; 树皮凋落高峰出现在高温干旱的8月; 松果凋落多在3和8月, 受降水影响明显。
凋落物是植物生长发育的产物, 其归还数量和季节动态首先受生物节律和物候期影响, 但同时也受到温度、水分等环境胁迫的影响(黄承才等, 2005)。因此, 凋落物动态常表现为一定的季节性和月际变动性。广西凭祥市马尾松Ⅰ类产区内生长良好的马尾松纯林的月凋落量变化表现为:10—11月最高, 12月至次年1月略低, 2—3月出现较大降幅, 持续降至8—9月的全年低谷期(周运超等, 2002)。本研究的马尾松纯林因遭受了酸雨危害, 其凋落量月变化明显与广西凭祥不同:5—6月凋落较少, 7月开始增多, 9—10月达到峰值, 其月凋落量呈现出随土壤湿度降低而增加的趋势。由相关分析得知, 月凋落动态对0~40 cm土层的土壤湿度变化的响应非常强烈(P<0.01), 这与当地受害马尾松林的根系多分布于0~40 cm土层(特别是腐殖质层和0~10 cm土层)有关(张治军等, 2008; 李志勇等, 2009)。长期土壤酸化造成的根系减少和分布浅层化, 尤其是起吸收作用的细根的死亡和退化, 间接导致了马尾松对土壤水分吸收能力的下降, 进而使地上部分尤其是枝叶的凋落对土壤干旱非常敏感。这是酸雨间接危害森林健康的一个重要途径。本研究还表明, 马尾松凋落动态对土壤湿度变化的响应存在时间滞后, 其详细的生理生态机制有待进一步研究。
冯宗炜. 2000. 中国酸雨对陆地生态系统的影响和防治对策[J]. 中国工程科学, 2(9): 5-11. |
黄承才, 葛滢, 朱锦茹, 等. 2005. 浙江省马尾松生态公益林凋落物及与群落特征关系[J]. 生态学报, 25(10): 2507-2513. DOI:10.3321/j.issn:1000-0933.2005.10.007 |
李志勇, 王彦辉. 2009. 酸化胁迫下的森林健康研究—以重庆铁山坪为例[M]. 北京: 中国农业出版社: 15-50.
|
卢立华, 贾宏炎, 何日明, 等. 2008. 南亚热带6种人工林凋落物的初步研究[J]. 林业科学研究, 21(3): 346-352. |
温远光, 黄承标, 梁宏温. 1987. 里骆林区杉木生态物候学的初步研究[J]. 生态学杂志, 6(6): 10-16. |
杨帆, 苗灵风, 胥晓, 等. 2007. 植物对干旱胁迫的响应研究进展[J]. 应用与环境生物学报, 13(4): 586-591. |
杨永森, 段雷, 靳腾, 等. 2006. 石灰石和菱镁矿对酸化森林土壤修复作用的研究[J]. 环境科学, 27(9): 1878-1883. |
张治军, 王彦辉, 袁玉欣, 等. 2006. 马尾松天然次生林生物量的结构与分布[J]. 河北农业大学学报, 29(5): 37-43. |
张治军, 王彦辉, 于澎涛, 等. 2008. 不同优势度马尾松的生物量及根系分布特征[J]. 南京林业大学学报:自然科学版, 32(4): 71-75. |
周运超, 张运吉, 谌红辉, 等. 2002. 施肥对马尾松人工中龄林生物归还的影响[J]. 南京林业大学学报:自然科学版, 26(4): 35-38. |
Meentemeyer V, Box E O, Thompson R. 1982. World patterns and amounts of terrestrial plant litter production[J]. Bioscience, 32(2): 125-128. DOI:10.2307/1308565 |