 2011, Vol. 47
2011, Vol. 47文章信息
- 田大伦, 杨超, 闫文德, 项文化, 方晰, 梁小翠
- Tian Dalun, Yang Chao, Yan Wende, Xiang Wenhua, Fang Xi, Liang Xiaocui
- 连栽杉木林不同生育阶段林下植被生物量
- Understory Vegetation Biomass in Successive Rotations of Different Aged Cunninghamia lanceolata Forests
- 林业科学, 2011, 47(5): 145-149.
- Scientia Silvae Sinicae, 2011, 47(5): 145-149.
- DOI: 10.11707/j.1001-7488.20110523
-
文章历史
- 收稿日期:2011-01-11
- 修回日期:2011-03-28
-
作者相关文章


2. 国家林业局 北京 100714;
3. 南方林业生态应用技术国家工程实验室 长沙 410004;
4. 国家野外科学观测研究站 会同 418307
2. State Forestry Administration Beijing 100714;
3. National Engineering Laboratory for Applied Technology of Forestry & Ecology in South China Changsha 410004;
4. National Field Station for Scientific Observation and Experiment Huitong 418307
林下植被是森林生态系统的一个重要组成部分,亦是森林生态系统中有机物质的生产者,在森林生态系统的物质循环研究中具有重要作用(Chapin,1983; Chastain et al., 2006),如维持森林生物多样性(褚建民等,2007)、提高人工林的水土保持功能(袁正科等,2002)和保护环境(刘苑秋等,2004; Fabia et al., 2002; Kume et al., 2003; Taylor et al., 2006)。对森林林下植被结构和生态功能的研究已有不少报道(Taylor et al., 2006; 吴鹏飞等,2008; 李春义等,2007; 段劼等,2010)。
杉木(Cunninghamia lanceolata)是我国亚热带特有树种,在我国森林资源中占有十分重要的地位。对杉木人工林林下植物的生态功能和作用的研究,自20世纪80年代就已经开始,如林下植被在杉木林生态系统养分循环中的作用(潘维俦等,1981; 冯宗炜等,1985; 方奇,1990),林下植被对杉木林地力和水土保持的影响(姚茂和等,1991a; 1991b; 张先仪等,1992; 盛炜彤等,1997; 盛炜彤,1986; 杨承栋等,1995),目前已取得不少成果。但从空间上一致和时间上连续地对杉木林林下植被动态特征进行的长期定位研究未见报道。
自1978年以来,对中南林业科技大学湖南会同杉木林生态系统国家野外科学观测研究站开展了杉木林结构和功能的系统定位研究(田大伦等,2003; 田大伦,2005a; 2005b; 方海波等,1998a; 1998b)。本研究对生态站6号集水区连栽杉木林不同生育阶段的林下植被种类及生物量动态特征进行了连续14年(1997—2010年)的定位研究,这对认识杉木林生态功能和养分平衡、提高林地地力及可持续经营具有十分重要的意义。
1 研究区概况研究区位于湖南会同杉木林生态系统国家野外科学观测研究站,国家林业局重点森林生态系统定位研究站———中南林业科技大学会同森林生态站(109°45'E,26°50'N)。该区属典型亚热带湿润性气候,年均气温16.8 ℃,年降水量1 100~1 400 mm,年均相对湿度80%以上。海拔300~500 m,为低山丘陵地貌。土壤为由震旦纪板溪系灰绿色板岩发育而来的山地黄壤。本次研究区设在生态站的6号集水区,其土壤pH为4.71±0.19,密度(1.31±0.02) g·cm-3,有机质含量(20.97±2.14) g·kg-1,全N含量(1.41±0.17)g·kg-1,全P含量(0.35±0.02) g·kg-1,全K含量(24.52±3.84) g·kg-1,全Ca含量(1.31±0.25) g·kg-1,全Mg含量(3.35±0.21) g·kg-1。
2 研究方法1989年冬季在生态站研究基地6号集水区,将1966年营造的杉木林进行皆伐,于1997年春季经炼山全垦整地后种植了杉木纯林,造林密度为2 700株·hm-2,造林后当年11月和翌年(1998年)5,8月对幼林进行了抚育,将杂草弃于林地,让其自行腐烂,以后未作其他处理。
在研究区山坡的上部、中部和下部共设置18个5 m × 5 m的样方,详细记录每个样方内杉木林林下植物种类,采用全挖法实测灌木和草本植物的鲜质量,分别将同种植物各取样本1.0 kg,在鼓风干燥烘箱内于80 ℃恒温下烘至恒质量,求出样本含水率。自1997年11月至2010年11月,于每年11月份测定1次连栽杉木林林下植被的生物量,共连续测定14年。
数据分析采用SPSS13.0统计软件处理。杉木林生育阶段按潘维俦等(1981)和吴中伦(1984)的原则划分。
3 结果与分析 3.1 幼林阶段林下植被生物量由表 3可见:3年生杉木幼林灌木层生物量为1 519.58 kg·hm-2,其中乔木树种盐肤木(Rhus chinensis)、大青(Clerodendrum cyrtophyllum)和泡桐(Paulownia)生物量共为885.68 kg·hm-2,占灌木层总生物量的58.28%,灌木植物杜茎山(Maesa japonica)和空心泡(Rubus rosaefolius)生物量共为366.33 kg·hm-2,占24.12%,藤本植物钩藤(Uncaria rhynchophylla)生物量为267.57 kg·hm-2,仅占17.60%;草本层生物量高达8 497.55 kg·hm-2,比灌木层多6 977.97 kg·hm-2,而且生物量在种类分布上极不均匀,芒草(Miscanthus sinensis)生物量为6 853.23 kg·hm-2,占草本层总生物量80.65%,而铜锤玉带草(Pratia nummularia)生物量为22.43 kg·hm-2,仅占0.27%。
|
|
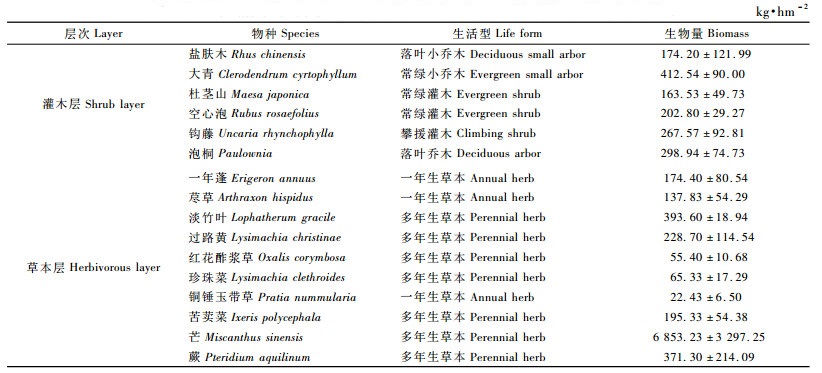
由表 4可见:当杉木林为5年生时,由于树冠扩展迅速,林下生态环境发生变化,林下植物生物量也随之产生变化,灌木植物生物量由3年生时的1 519.58 kg·hm-2,下降至601.51 kg·hm-2,减少了918.07 kg·hm-2; 草本植物生物量由3年生时的8 497.55 kg·hm-2,下降至683.09 kg·hm-2,减少了7 814.46 kg·hm-2。
|
|
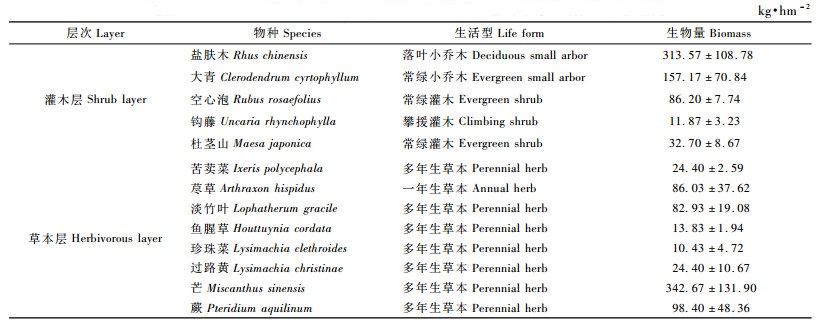
由表 5可见:9年生杉木林灌木层生物量为2 594.48 kg·hm-2,以落叶小乔木类植物为主体,生物量为1 967.04 kg·hm-2,占灌木层生物量的75.82%,灌木类植物生物量为627.44 kg·hm-2,只占24.18%;草本层生物量为2 658.45 kg·hm-2,以芒草和蕨类植物占优势,生物量达2 097.01 kg·hm-2,占草本层生物量的78.89%。
|
|
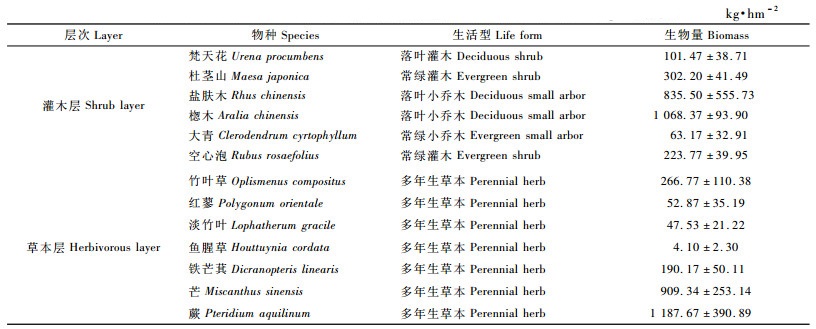
由表 6可见:10年生杉木林林下植被生物量为6 646.02 kg· hm-2; 灌木层生物量为4 148.58 kg·hm-2,占总生物量的62.4%,其中楤木(Aralia chinensis)生物量为2 886.01 kg·hm-2,占灌木层生物量的69.57%;草本层生物量为2 497.44 kg·hm-2,其中蕨类植物生物量为1 856.51 kg·hm-2,占草本层生物量的74.34%,淡竹叶(Lophatherum gracile)生物量为32.80 kg·hm-2,仅占1.31%。
|
|
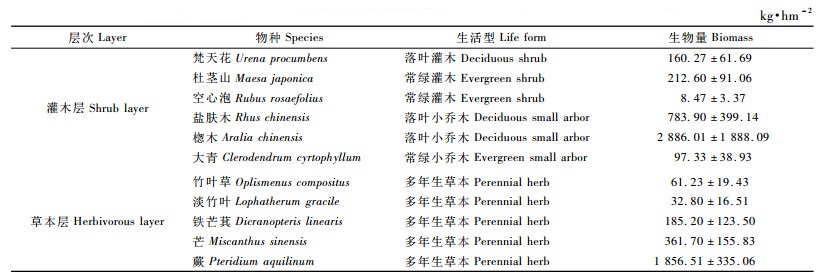
当杉木林迈入杆材阶段,自然整枝仍很强烈,又为自然稀疏剧烈时期,但是更为林分材积迅速增长时期(吴中伦,1984),杉木不仅占据了空间位置,而且还改变了土壤养分的可利用性,从而影响到林下植被生物量及其组成与分配。由表 7可见:13年生杉木林林下灌木层生物量为1 267.81 kg·hm-2,比10年生林分林下灌木层生物量减少了2 880.77 kg·hm-2,盐肤木在生物量组成中占绝对优势; 草本层生物量比10年生林分林下草本层生物量减少了1 789.67 kg·hm-2,生物量组成以铁芒萁(Dicranopteris linearis)和亚大苔草(Carex brownii)为主。
|
|
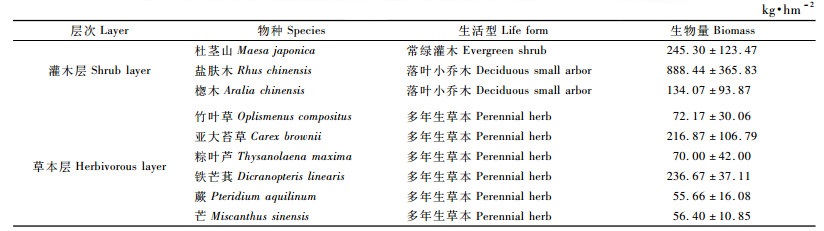
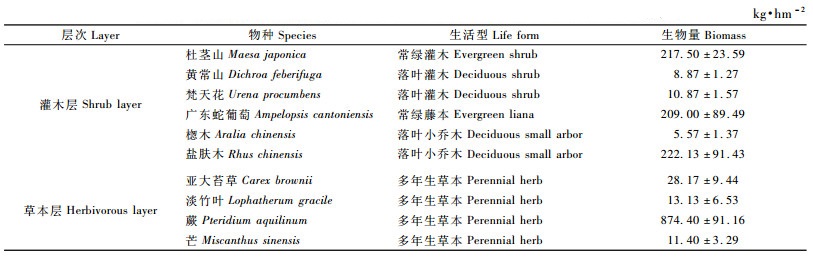
由表 8可见:14年生杉木林下灌木层生物量为673.94 kg · hm-2,比13年生林分减少593.87 kg·hm-2,生物量组成以杜茎山(占32.27%)、盐肤木(占32.96%)和广东蛇葡萄(Ampelopsis cantoniensis)(占31.01%)为主; 草本层生物量为927.10 kg·hm-2,比13年生林分增加219.33 kg·hm-2,以蕨类植物的生物量最高,为874.40 kg·hm-2,占草本层总生物量的94.31%。
|
|
杉木林在14年的生长过程中,林下植被生物量以林分未郁闭前的幼林阶段3年生时最大,达10 017.13 kg·hm-2,其中灌木植物为1 519.58 kg·hm-2,草本植物为8 497.55 kg·hm-2,草本植物生物量为灌木植物生物量的5.6倍,占地被物总生物量的84.8%。当杉木林进入第5年生时,由于杉木树冠的扩展,林分郁闭度加大,林下光照减弱,地被物生物量由10 017.13 kg·hm-2,下降至1 284.60 kg·hm-2,下降了87.2%,尤以草本层生物量下降率(92.0%)明显。杉木林进入速生阶段,林分的自然整枝开始,林冠相对缩短,林冠层逐步上升(吴中伦,1984; Chastain et al., 2006),与此同时,林内光照加强,环境条件改善,使林下植被生物量得到增加,林分为9年生时,林下植被生物量为5 252.93 kg·hm-2,比5年生时增加75.5%,林分为10年生时,林下植被生物量为6 646.02 kg·hm-2,又比9年生时增加21.0%。林分进入杆材阶段,虽然自然整枝强烈,但由于林冠相接和杉木自身需要占据林地空间加大,林下植被对林地空间的争夺能力减小,而伴随着生物量减少,当林分为13年生时,林下植被生物量为1 975.58 kg·hm-2,比10年生林分减少70.3%,林分14年生时,林下植被生物量达最低值,为1 601.04 kg·hm-2,又比林分13年生时减少19.0%。连栽杉木林从幼林阶段到杆材阶段的14年生长过程中,林下植被生物量表现出波动性的下降趋势。不同生育阶段杉木林林下植被生物量受林分郁闭度和林龄的影响较大,这与刘煊章等(1997)和闫文德等(2003)对会同第二代杉木人工林林下植被生物量的研究结果一致。
本研究仅针对会同连栽杉木林幼林阶段到杆材阶段(1997—2010年)林下植被生物量的动态变化,而成熟与过熟阶段至采伐利用期的林下植被生物量的动态研究,仍需继续进行定位监测。
褚建民, 卢琦, 崔向慧, 等. 2007. 人工林林下植被多样性研究进展[J]. 世界林业研究, 20(3): 9-13. |
段劼, 马履一, 贾黎明, 等. 2010. 抚育间伐对侧柏人工林及林下植被生长的影响[J]. 生态学报, 30(6): 1431-1441. |
方海波, 田大伦, 康文星. 1998a. 杉木人工林间伐后林下植被生物量的研究[J]. 中南林学院学报, 18(1): 5-9. |
方海波, 田大伦, 康文星. 1998b. 杉木人工林间伐后林下植被养分动态的研究[J]. 中南林学院学报, 18(2): 1-5. |
方奇. 1990. 加强土壤和地被物管理对杉木生态系统生物量能量利用与养分循环的影响[J]. 林业科学, 26(3): 201-208. |
冯宗炜, 陈楚莹, 王开平, 等. 1985. 亚热带杉木纯林生态系统中营养元素的积累、分配和循环的研究[J]. 植物生态学与地植物学丛刊, 9(4): 245-255. |
李春义, 马履一, 王希群, 等. 2007. 抚育间伐对北京山区侧柏人工林林下植物多样性的短期影响[J]. 北京林业大学学报, 29(3): 60-66. |
刘煊章, 田大伦, 文仕知, 等. 1997. 第二代杉木人工林林下地被物生物量和养分积累的定位研究Ⅰ.林下地被物生物量动态[J]. 林业科学, 33(增刊2): 19-25. |
刘苑秋, 罗良兴, 杨国平, 等. 2004. 退化红壤重建森林林下植被恢复及其环境影响分析[J]. 江西农业大学学报, 26(5): 695-699. |
潘维俦, 田大伦, 李利村, 等. 1981. 杉木人工林养分循环的研究(一)不同生育阶段杉木林的产量结构和养分动态[J]. 中南林学院学报, 1(1): 1-21. |
盛炜彤, 杨承栋. 1997. 关于杉木林下植被对改良土壤性质效用的研究[J]. 生态学报, 17(4): 377-385. |
盛炜彤. 1986. 关于提高杉木人工林生产力的几个问题[J]. 浙江林业科技, 6(1): 9-15. |
田大伦. 2005a. 杉木林生态系统功能过程[M]. 北京: 科学出版社.
|
田大伦. 2005b. 杉木林生态系统研究方法[M]. 北京: 科学出版社.
|
田大伦, 康文星, 文仕知. 2003. 杉木林生态系统学[M]. 北京: 科学出版社.
|
吴鹏飞, 朱波. 2008. 桤柏混交林林下植被结构及生物量动态[J]. 水土保持通报, 28(3): 44-48. |
吴中伦. 1984. 杉木[M]. 北京: 中国林业出版社: 162-163.
|
闫文德, 田大伦, 焦秀梅. 2003. 会同第二代杉木人工林林下植被生物量分布及动态[J]. 林业科学研究, 16(3): 323-327. |
杨承栋, 焦如珍, 屠星南, 等. 1995. 发育林下植被是恢复杉木人工林地力的重要途径[J]. 林业科学, 31(3): 275-283. |
姚茂和, 盛炜彤, 熊有强. 1991a. 林下植被对杉木林地力影响的研究[J]. 林业科学研究, 4(3): 246-252. |
姚茂和, 盛炜彤, 熊有强. 1991b. 杉木林林下植被及其生物量的研究[J]. 林业科学, 27(6): 644-648. |
袁正科, 田育新, 李锡泉, 等. 2002. 缓坡梯土幼林林下植被覆盖与水土流失[J]. 中南林学院学报, (2): 21-24. |
张先仪, 邓宗付, 李旭明. 1992. 间伐杉木林下植被演替和水土保持影响的研究[M]. 北京: 中国科学技术出版社: 168-180.
|
Chapin F S. 1983. Nitrogen and phosphorus nutrition and nutrition cycling by evergreen and deciduous understory shrubs in an Alaskan black spruce forests[J]. Canadian Journal of Forest Research, 13(5): 773-781. DOI:10.1139/x83-107 |
Chastain R A, Currie W S, Townsend P A. 2006. Carbon sequestration and nutrient cycling implications of the evergreen understory layer in Appalachian forests[J]. Forest Ecology and Management, 231: 63-77. DOI:10.1016/j.foreco.2006.04.040 |
Fabia A, Martins M C, Cerverira C. 2002. Influence of soil and organic residue management on biomass and in a Eucalyptus globules labill plantation[J]. Forest Ecology and Management, 171: 87-100. DOI:10.1016/S0378-1127(02)00463-2 |
Kume A, Satomura T, Tsubei N. 2003. Effects of understory vegetation on the ecophysiological characteristics of and overstory pine, Pinus densiflora[J]. Forest Ecology and Management, 176: 195-203. DOI:10.1016/S0378-1127(02)00282-7 |
Taylor A H, Jang S W, Zhao L J. 2006. Regeneration patterns and tree species coexistence in old growth Abies-Picca forests in southwestern China[J]. Forest Ecology and Management, 223: 303-317. DOI:10.1016/j.foreco.2005.11.010 |