 2011, Vol. 47
2011, Vol. 47文章信息
- 韩宗先, 王维, 胡锦矗
- Han Zongxian, Wang Wei, Hu Jinchu
- 重庆金佛山黑叶猴的春季生境选择
- Habitat Selection by Francois' Langur in Jinfo Mountain in Spring
- 林业科学, 2011, 47(4): 121-128.
- Scientia Silvae Sinicae, 2011, 47(4): 121-128.
-
文章历史
- 收稿日期:2009-10-19
- 修回日期:2010-03-21
-
作者相关文章


2. 重庆市林业局 重庆 400015;
3. 西华师范大学珍稀动植物研究所 南充 637002
2. Chongqing Forestry Bureau Chongqing 400015;
3. Institute of Rare Animals and Plants, China West Normal University Nanchong 637002
黑叶猴(Trachypithecus francoisi francoisi)在全球范围内分布很窄,仅分布于越南北部、中国的广西、贵州和重庆(叶智彰,1993;王应祥等,1999),是我国Ⅰ级保护动物,东南亚特产种,IUCN红皮书将其列为易危种(VU),中国濒危动物红皮书将其列为濒危种(E)。一个时期以来,由于森林资源的破坏,黑叶猴栖息地破碎化加剧,栖息地丧失越来越严重,黑叶猴资源数量不断下降(吴名川等,1987;黄乘明等,2006)。作为一种珍稀濒危动物,黑叶猴在分类、系统进化、生态学、行为学等领域具有十分重要的研究价值(叶智彰,1993;王应祥等,1999)。目前未见国外的研究报道,国内对野生黑叶猴的研究主要集中在资源调查(李明晶等, 1989;李明晶,1994;张含藻等,1992; 1994;刘万福等,1995;唐洪通等, 1998;苏化龙等,2002)、日活动行为类型(黄中豪等,2007)、活动节律(黄乘明等,2006;周岐海等,2007a)、活动时间分配(Huang et al., 2003;罗杨等,2005; 2007;周岐海等,2001;2009;黄乘明等,2007)、家域(陈正仁等,2001)、食性(罗杨等,2000;李友邦等,2008;Zhou et al., 2006)等方面,对生境利用的研究仅见周岐海等(2007b)的报道。从研究地点来看,主要集中在贵州麻阳河、广西扶绥和弄岗。重庆是黑叶猴分布的北缘,生态环境条件与贵州和广西有所不同,但目前尚无重庆地区黑叶猴生态学的研究报道,因此在金佛山自然保护区开展黑叶猴研究,以期为有效保护黑叶猴资源,制订合理的保护方案提供科学依据。
1 自然概况金佛山国家级自然保护区位于107°00′—107°20′E,28°50′—29°20′N,总面积418.5 km2,行政区划属重庆南川市。金佛山属大娄山脉,为石灰岩喀斯特地貌,最高海拔2 251 m,最低海拔600 m。该区属亚热带湿润季风气候,温和多雨,冬微寒夏暖,四季分明。整个山体四周有多层次的常流瀑布,形成了数十条溪流,为多种动植物生长繁衍提供了良好的条件(马建伦等,2007)。
黑叶猴分布区位于金佛山东北侧的狭谷地带,总面积约50 km2。该区年平均气温12.5℃,相对湿度80%~90%,无霜期190~280天,年降雨量1 395 mm。黑叶猴生活于沟谷的悬崖陡壁处,植被繁茂的乔灌混交林,食物种类多,植被群落复杂,主要有桑科、山毛榉科、山茶科、杜鹃科等30余科,300多种组成的植物群落。黑叶猴生活区海拔悬殊较大,最低600 m,最高1 800 m。随着气候及食物的变化,黑叶猴在不同海拔作季节性迁移(唐洪通等,1998)。
本研究区位于庙坝村吊咀(图 1),面积约20 km2,海拔980~1 650 m。研究区内多数是植被繁茂的乔灌混交林,仅有少数区域是灌木从; 植被群落复杂,主要有桑科、樟科、山茶科、杜鹃花科、壳斗科等的多种植物,因而黑叶猴食物丰富。黑叶猴常在海拔1 290~1 400 m的峭壁石岩上或石岩腔下过夜,其夜宿地既不受风雨袭击,天敌又不易到达。夜宿地上下、左右均是茂密的森林,采食十分方便,通常夜宿地下面便是小溪,饮水也较为方便。

|
图 1 研究区域示意 Figure 1 The schematic diagram of study area |
应用资源选择指数(resource selection index, RSI)(Manly et al., 1993;Lechowicz,1982)和资源选择函数(resource selection functions, RSFs)(Boyce et al., 1999;Lennon,1999)研究黑叶猴对生境的选择。
2.1 资源选择函数(RSFs)的含义资源选择函数在研究动物的生境偏好方面具有明显优势,它用已利用资源与可利用资源的比率来计算(Boyce et al., 1999;韩宗先等,2004)。具体计算方法如下。
对于生境中的一种资源i,物种对它的选择率为:ωi=οi/πi,其中οi是资源i中被使用的比例,πi=ai/a+,a+是所有可供使用的资源单位,ai是其中资源i可以被使用的单位。
由于物种对生境的选择往往受食物、遮蔽物和水热条件等多种因素的制约,所以资源选择函数一般表现为一个包括多个独立生境变量的线性对数模型:ω(χ)=exp(β0+β1χ1+β2χ2+…+βkχk), 其中χ代表了不同的独立生境变量,β表示选择系数。那么,物种对生境的选择概率为:m(χ)=exp(β0+β1χ1+β2χ2+…+βkχk)/[1+exp(β0+β1χ1+β2χ2+…+βkχk)], 当m(χ)的取值为1或0时,即表示选择或不选择时,选择系数β可以由逻辑斯蒂回归系数来估计(Boyce et al., 1999)。
逻辑斯蒂回归在1967年首次用于多变量分析(Hosmer et al., 1989),现在它是研究二值响应变量(例如,有和无)或有序响应变量与一组自变量之间关系的一种标准统计方法,目前在大多数统计软件(如SPSS,SAS等)中都有固定的模块来实现(张文彤,2002)。
2.2 资源选择指数的计算方法 2.2.1 资源选择率ωi对于资源i,物种对它的选择率为:ωi=οi/πi,πi=ai/a+,其中οi, ai, a+的定义同前。
2.2.2 资源选择系数Wi和资源选择指数EiWi=ωi/∑ωi, ωi为资源选择率; Ei=(Wi-1/n)/(Wi+1/n), n为某类资源的等级数。Ei值介于-1和+1之间,若Ei>0.1表示喜爱,Ei=1表示特别喜爱,Ei=0为随机选择,-0.1<Ei<0.1表示几乎随机选择,Ei<-0.1表示不喜爱,Ei=-1为不选择。
2.3 样方的设置 2.3.1 已利用生境样方的设置以猴群活动时间超过30 min处为中心,设立50 m×50 m正方形大样方30个,在大样方中间设立2个互相垂直的10 m×50 m的长方形样方,这样中间交叉得到一个10 m×10 m的小正方形样方,四角就是4个20 m×20 m中型正方形样方。各生境变量的测定方法如下:
郁闭度:整个50 m×50 m大样方的郁闭度, 共分为4级, 即 < 30%, 30%~50%, 50%~80%, >80%。
坡度:整个50 m×50 m大样方所处地坡度,共分3级, 即 < 30°, 30°~60°, >60°。
坡向:整个50 m×50 m大样方所处地的坡向,共分4级, 即东坡(45°~ 135°)、南坡(135°~225°)、西坡(225°~315°)、北坡(315°~45°)。
坡位:整个50 m×50 m大样方所处地的坡位, 分为3级,即坡上位(山岗或坡上部)、坡中位(山腰或坡中部)、坡下位(山谷或坡下部)。
海拔:10 m×10 m小样方中心的海拔(m)。
植被:以植被的生长型外貌而定,分为乔木林、乔木-灌木林、灌木丛。乔木林:整个50 m×50 m样方内有乔木,但没有灌木(在本文中“灌木”是指胸径大于5 cm的灌木以及高度小于5 m且胸径大于5 cm的乔木幼树)。乔木-灌木林:整个50 m×50 m样方内既有乔木,又有灌木。灌木丛:整个50 m×50 m样方内有灌木,但没有乔木。
人为干扰:以50 m×50 m大样方中心离水力发电站、地质钻探点、开矿点、林间小路、山民采药、放牧等人为活动点的直线距离为划分标准, 大于1 000 m为轻度干扰, 500 m~1 000 m为中度干扰, 小于500 m为重度干扰。
水源:根据泉水、溪流等到调查样方中心的直线距离划分为3级, 小于200 m为近水源, 200 m~500 m为中水源, 大于500 m为远水源。
乔木密度:从4个20 m×20 m正方形样方中测得的乔木平均数量[株·(400 m2)-1]。
乔木胸径:4个20 m×20 m正方形样方中,离中心点最近的一棵乔木树的胸径的平均值(cm)。
灌木密度:从4个20 m×20 m正方形样方中测得的灌木平均数量[株·(400 m2)-1]。
灌木胸径:4个20 m×20 m正方形样方中,离中心点最近的一棵灌木树的胸径的平均值(cm)。
洞穴距离:以10 m×10 m正方形样方中心点到最近的洞穴的直线距离(m),分为3级,即 < 200 m, 200 m~500 m, >500 m。
食物丰富度:50 m×50 m大样方中可供使用的食物数量,分为充足、基本满足、贫乏3个等级。统计黑叶猴春季喜食的灯台树(Cornus controversa)、香叶树(Lindera communis)、毛叶木姜子(Litsea mollis)、山梨(Pyrus ussuriensis)、构树(Broussonetia papyrifera)、木姜子(Litsea wilsoni)、山樱花(Cerasus serrulata)、山桃(Amygdalus dawidiana)、山桑(Morus mongolica)和野桐(Mallotus japonicus var. floccosus)的株数,若大于20株,记为充足, 10~20株记为基本满足, 小于10株记为贫乏。
2.3.2 对照样方的设置为保证对照样方的随机性, 采用系统样方方格抽样法(张洪海等,1999;韩宗先等,2004), 在研究区域内设置30个对照样方, 测定了同样的生境参数。具体作法为,在研究区域内海拔每上升100 m设一条样线,在每条样线上每隔500 m设置一个对照样方(里面同样设立小样方),使对照样方的抽取面积基本覆盖整个研究区域。
2.4 数据处理数据的分析处理采用SPSS11.0。
3 结果与分析 3.1 黑叶猴对生境资源的选择率ωi、选择系数Wi和选择指数Ei黑叶猴对各种生境资源的选择率、选择系数和选择指数见表 1。
|
|
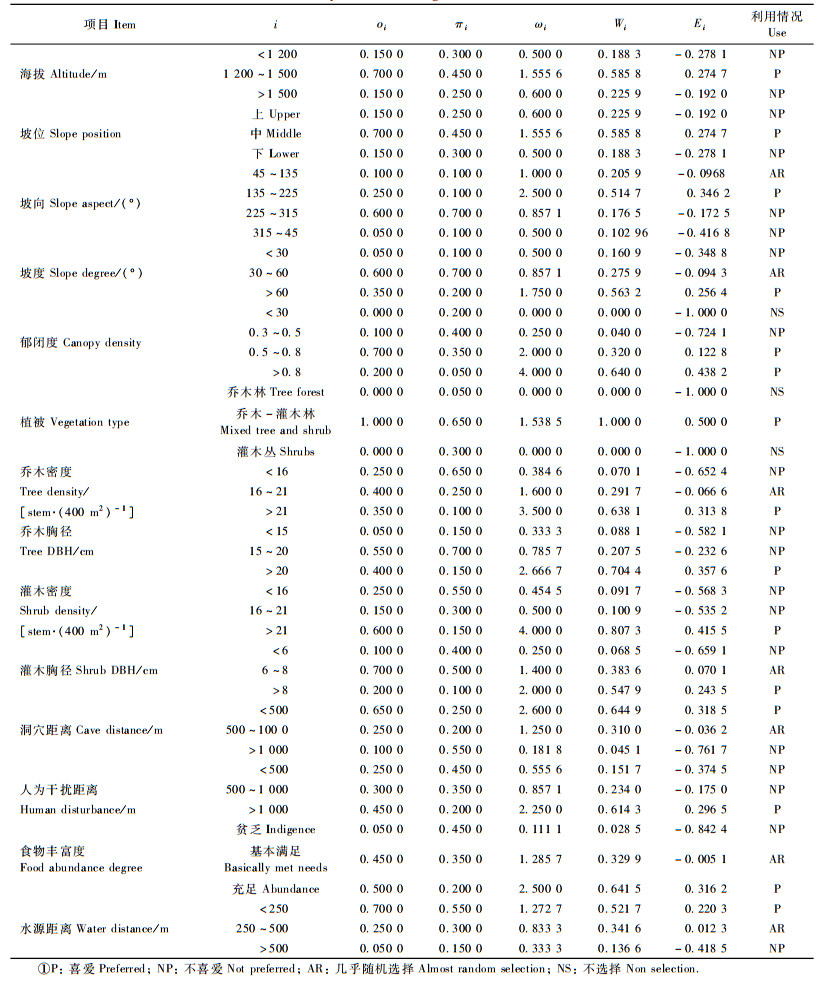
资源选择指数的统计结果表明,对于海拔,黑叶猴喜爱海拔1 200~1 500 m的生境,不喜爱海拔低于1 200 m和高于1 500 m的生境。对于坡位,黑叶猴喜爱选择坡中位,不喜爱选择坡上位和坡下位。对于坡向,黑叶猴喜爱南坡,几乎随机选择东坡,不喜爱选择西坡和北坡。对于坡度,黑叶猴喜爱大于60°的生境,不喜爱小于30°的生境,对于30°~60°的生境几乎随机选择。对于郁闭度,黑叶猴喜欢郁闭度大于0.5的生境,不喜爱选择郁闭度0.3~0.5的生境,不选择郁闭度小于0.3的生境。对于植被,黑叶猴喜爱选择乔木-灌木混交林,不选择只有单一乔木或者是灌木的生境。对于乔木密度,黑叶猴喜爱乔木密度大于21株·(400 m2)-1的生境,不喜爱选择乔木密度小于16株·(400m2)-1的生境,几乎随机选择乔木密度在16~21株·(400m2)-1的生境。对于乔木大小,黑叶猴喜爱乔木胸径大于20 cm的生境,不喜爱选择乔木胸径等于和小于15 cm的生境。对于灌木密度,黑叶猴不喜爱灌木密度小于和等于21株·(400m2)-1的生境,喜爱灌木密度大于21株·(400m2)-1的生境。对于灌木大小,黑叶猴喜爱灌木胸径大于8 cm的生境,对于灌木胸径6~8 cm的生境随机选择,不喜爱选择小于6 cm的生境。对于洞穴距离,黑叶猴喜爱洞穴距离小于500 m的生境,不喜爱洞穴距离大于1 000 m的生境,几乎随机选择500~1 000 m的生境。对于人为干扰,黑叶猴喜爱在距离人为干扰大于1 000 m的生境活动,不喜爱选择人为干扰距离小于和等于1 000 m的生境。对于食物丰富度,黑叶猴喜爱食物充足的生境,几乎随机选择基本满足的生境,不喜爱食物贫乏的生境。对于水源,黑叶猴喜爱水源距离小于250 m的生境,不喜欢水源距离大于500 m的生境,几乎随机选择水源距离在250~500 m的生境。可见,黑叶猴春季最适生境为郁闭度大于50%、乔木和灌木密度大于21株·(400m2)-1、灌木胸径大于8 cm、人为干扰大于1 000 m、洞穴距离小于500 m、水源距离小于250 m、南坡的乔木-灌木混交林。
从表 1可以看出,资源选择指数只能比较黑叶猴对同一资源项目中不同资源等级的偏爱程度,不能判断哪些资源项目起主导作用, 而资源选择函数正好弥补了这一缺陷。
3.2 黑叶猴的资源选择函数应用逻辑斯蒂回归拟合资源选择函数变量的选择系数是以变量相互独立为前提的(Boyce et al., 1999;Lennon,1999), 故对14个生境因子进行了相关分析。在变量两两比较所得到的91个相关系数中,绝对值大于0.5且有统计学意义的有4个(表 2)。考虑到生境因子的独立性和代表性, 从14个因子中筛选出了11个进行逻辑斯蒂回归, 即:海拔、坡向、植被、上层乔木郁闭度(CA)、乔木胸径、乔木密度、灌木胸径(DBH)、灌木密度、洞穴距离(DC)、水源距离、人为干扰距离。
|
|

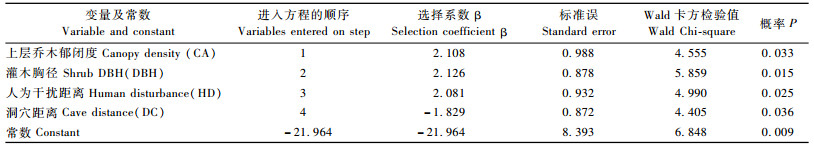
把所有参数标准化后, 采用Forward/Conditional (以假定参数为基础作似然比概率检验, 向前逐步选择自变量)法进行逻辑斯蒂回归分析, 最终进入函数方程的有明显统计学意义的变量为,郁闭度、灌木胸径、人为干扰、洞穴距离(表 3)。故黑叶猴的生境资源选择函数为:logit (P)=-21.964+2.126×DBH+ 2.108×CA+ 2. 081×HD-1.829×DC。
|
|
根据拟合出的资源选择函数, 黑叶猴对生境的选择概率为:P=elogit (P)/(1+elogit(P)),模型的正确预测率可达87.5%。
从以上分析可以看出,影响黑叶猴生境选择的关键因子有4个,根据选择系数的绝对值大小及采用Enter法(所有自变量强制进入回归方程)拟合的结果,这4个因子依其重要性可排序为:上层乔木郁闭度、灌木胸径、人为干扰距离、洞穴距离。
4 讨论野生动物总是生活在特定的生境中,生境作为物种生存的空间,为物种提供了包括食物、水源、隐蔽条件和繁殖场所等资源条件(Ables,1969;Baily,1984;马建章等,2004;蒋志刚,2004)。生境决定了可获得或可利用资源的数量和质量,直接或间接决定着种内和种间竞争的强度,对物种持续生存繁衍有着重大影响,是推动物种发展进化的重要的生态因素(尚玉昌,1998;魏辅文等,1998;孙儒泳,2001;李言等,2008)。由于不同生态因子对特定物种生存意义的差异在空间分布上的普遍异质性,动物倾向于选择具有最佳生态因子的组合,以保证最大的适合度(尚玉昌,1998;颜忠诚等,1998)。在具体的表现上,不同生态因子对物种生境选择有不同的影响(尚玉昌,1998;孙儒泳,2001;McLoughlin et al., 2002)。在金佛山自然保护区,上层乔木郁闭度、灌木胸径、人为干扰距离、洞穴距离对黑叶猴生境选择有重要影响。
黑叶猴为半树栖半地栖的灵长类(黄乘明等,2006),除夜晚在峭壁洞穴中栖息,白昼多数时间在树上活动,这就要求森林较繁茂。上层乔木郁闭度是反映森林繁茂与否以及森林质量的一个综合指标。郁闭度较大的生境不仅为黑叶猴提供了丰富的食物资源,而且提供了良好的隐蔽条件。在本研究中,食物丰富度与上层乔木郁闭度密切相关(相关系数为0.609),上层乔木郁闭度的大小,在较大程度上代表了食物资源的丰富度。黑叶猴在高大的树木上更容易扫描周围环境并及时发现食物和潜在的危险。食物是动物生命物质和能量的来源,是动物生存、繁衍和进化的基础(孙儒泳,2001)。对于绝大多数动物来说,获取食物和躲避捕食者是它们存活策略的主要组成部分(Lima et al., 1990)。越来越多的研究表明,灵长类对栖息地的选择与食物资源状况(Olupot et al., 1997;Kaplin, 2001;Wu et al., 2004)和捕食风险(Cowlishaw,1997;Enstam et al., 2004)有密切的关系。据Cowlishaw(1997)和Enstam等(2004)研究,灵长类更喜欢选择那些能够提供更多食物、更多避难所或更多保护性的遮盖物或更容易发现捕食者的栖息地,本研究也与此相符。黑叶猴喜爱选择上层乔木郁闭度较大(>0.5)的生境,这种生境不仅食物资源丰富,而且提供了良好的隐蔽条件,并且有较多的高大树木,在高大树木上黑叶猴更容易扫描周围环境并及时发现食物和潜在的危险。据刘瑛等(2009)和黎大勇等(2006)报道,川金丝猴(Rhinopithecus roxellana)和滇金丝猴(R.bieti)生境适宜性的指示指标主要是食物要素、隐蔽条件要素和人为干扰要素等,郁闭度较好地反映了食物要素和隐蔽条件要素,滇金丝猴常在郁闭度较高(40%~80%)的树林活动,郁闭度较低的树林(20%~40%)则很少光顾,郁闭度<20%的基本不去。这与本研究的结论一致。
灌木大小影响着黑叶猴对生境的选择。黑叶猴在森林中移动和漫游时,常常要借助灌木搭桥,林中灌木为黑叶猴移动和漫游提供了通道。所以,黑叶猴喜爱选择灌木胸径较大的乔木-灌木混合林。若灌木胸径太小,不能承受其体重,将会坠地。据刘璐(2009)研究,宽阔水国家级自然保护区黑叶猴首选乔木-灌木混合林,常在高灌层和灌木丛活动,在高灌层和灌木丛活动时间占60.05%。据张鹏等(2006)报道,秦岭川金丝猴53%的时间在低植被层(low stratum)活动,亚成年雌性和青少年猴更加经常在低植被层(灌木层和树叉处)活动。据张强等(2008)研究,藏酋猴常在3 m以上的灌木层处活动,很少在l~2 m和1 m以下灌木层处活动。灌木层不仅是黑叶猴空间利用的重要层片,同时也提供食物资源(罗杨等,2000;Zhou et al., 2006)。在金佛山自然保护区,香叶树、毛叶木姜子、山樱花、山桃、山桑、沙柳(Salix psammophila)、杜鹃(Rhododendron simsii)和石灰花楸(Sorbus folgneri)等灌木都是黑叶猴的食物。
外来干扰可作用于动物的生境选择和行为,进而对种群未来产生深远影响(马建章等,2004;蒋志刚,2004;黎大勇等,2006)。现在许多珍稀野生动物的活动区域明显缩小,主要原因就是人为干扰(马建章等,2004;蒋志刚,2004)。众多研究(黄乘明等,2000;丁伟等,2003;顾志宏等,2007;周岐海等,2007a;2007b;黄勇等,2008;周汝良等,2008;刘瑛等,2009)表明,人类活动干扰导致灵长类适宜生境丧失、生境破碎化,严重影响灵长类的生境选择。本研究区域周边住有少量农家,且有一条公路贯穿,因而人为干扰成为影响黑叶猴生境选择的关键因子之一。周岐海(2007b)证实,在广西扶绥珍稀动物保护区,黑叶猴更喜爱选择在山腰觅食活动,而较少选择山脚,原因是山脚是人为干扰最大的地方,山腰人为干扰较小。据黎大勇等(2006)报道,滇金丝猴对人类的回避表现明显,猴群常常选择远离人群的生境,一般离人类活动频繁地很远(>1 000 m)。
黑叶猴通常利用悬崖峭壁上的石洞作为夜宿地(吴名川等,1987;Cyril et al., 2006;周岐海等,2007b;郭宝华等,2009)。当猴群在某个区域觅食时,它们通常在这一区域内或附近的某个固定洞穴过夜(周岐海等,2004;郭宝华等,2009)。因此,洞穴的多少和距离明显影响黑叶猴的生境选择。黑叶猴的日移动距离不大(295~1 015 m)(周岐海等,2004),故喜爱选择洞穴上下、左右均是采食场的茂密森林,这样采食十分方便,有利于节省能量(周岐海等,2007;郭宝华等,2009)。已有研究证实,洞穴的位置和可利用性是影响灵长类栖息地利用的因素之一(Harrison,1983;Chapman et al., 1989;Boonratana,2000;周岐海等,2007b)。例如,Chapman等(1989)对蜘蛛猴(Ateles geoffroyi)的研究发现,它们倾向于选择离夜宿地较近的生境采食,这有利于节省时间和能量。周岐海(2007b)研究证实,在广西扶绥珍稀动物保护区,黑叶猴喜爱选择有夜宿岩洞或靠近夜宿岩洞的生境觅食,在A、C、D、E 4个觅食斑块内,有绝大部分夜宿时间利用的4个岩洞。非人灵长类选择有天然岩洞或靠近天然岩洞的生境活动,与安全因素有关(McGrew et al., 2003;Cyril et al., 2006),当遇到捕食袭击等危险情况时,可很快返回到洞穴,脱离危险。
水是动物生活所必需的物质资源,也是动物最重要的生存条件之一(孙儒泳,2001)。据谢东明等(2009)研究,太行山区的水资源相对缺乏,地表径流极小,降水偏少,冬季尤甚。太行山猕猴选择离水源近(< 1 000 m)的地方活动,直接饮水可能是猕猴在干旱冬季获得足量水分的重要途径。黑叶猴的生活区为喀斯特山区,整个山体有多层次的常流瀑布,构成了数十条溪流,因而水资源丰富,加之春季主要以灯台树、香叶树、毛叶木姜子、山梨、构树、木姜子、山樱花、山桃、山桑和野桐等植物的叶(嫩叶和成熟叶)为食,食物水分含量较高,故水源不是影响黑叶猴生境选择的关键因子。据黎大勇等(2006)和Su等(1998)报道,水源因子不是影响川金丝猴和滇金丝猴的生境选择的关键因子。另据Suzuki(1965)和Wada(1964)报道,日本猴(Macaca fuscata)经常在温泉附近活动可能主要是为了取暖,而不是饮水。以上的研究结果与其研究对象所处的环境和气候有密切关系,川金丝猴和滇金丝猴活动的地区降水较多,水资源丰富较丰富,植被茂盛,食物充足,猴子很容易从食物和环境中获取足够的水分; 日本猴活动地区冬季降雪较多,猴子容易从野外补充水分。
生境选择是野生动物对栖息地的非随机利用,影响个体生存力和适合度等一系列行为反应(Block et al., 1993;尚玉昌,1998;孙儒泳,2001;罗振华等,2008)。为长期有效地保护这一珍稀濒危物种,应当重点加强对黑叶猴最适生境乔木-灌木混交林的保护; 在缺少乔木树的灌木丛中,栽植一些黑叶猴喜食的乔木植物; 严格控制当地村民进山挖药、挖兰花、采笋、放牧等活动,尽可能减少人为活动对黑叶猴生境的破坏和影响。同时,在当地实施退耕还林政策,将坡耕地退还种植林木时,应考虑生态效益,种植一些乡土树种和黑叶猴喜食的植物种类,如灯台树、香叶树、毛叶木姜子、山梨、构树、木姜子、山樱花、山桃、山桑、野桐、沙柳等。
陈正仁, 罗杨, 汪双喜. 2001. 贵州麻阳河黑叶猴家域及影响因素初探[J]. 贵州林业科技, 29(2): 34-37. |
丁伟, 杨士剑, 刘泽华. 2003. 生境破碎化对黑白仰鼻猴种群数量的影响[J]. 人类学学报, 22(4): 338-344. |
顾志宏, 金昆, 刘世荣, 等. 2007. 四川省白河自然保护区川金丝猴生境评价[J]. 林业科学, 43(8): 96-99. |
郭宝华, 张明海, 李迪强, 等. 2009. 贵州麻阳河国家级自然保护区黑叶猴夜宿地选择的研究[J]. 野生动物杂志, 30(2): 64-69. |
韩宗先, 魏辅文, 张泽钧, 等. 2004. 蜂桶寨自然保护区小熊猫对生境的选择[J]. 兽类学报, 24(3): 185-192. |
黄乘明, 周岐海, 李友邦, 等. 2006. 广西扶绥黑叶猴活动节律和日活动分配[J]. 兽类学报, 26(4): 380-386. |
黄乘明, 韦显盛, 周岐海, 等. 2007. 栖息地质量对黑叶猴活动时间分配的影响[J]. 兽类学报, 27(4): 338-343. |
黄中豪, 周岐海, 李友邦, 等. 2007. 弄岗黑叶猴的日活动类型和活动时间分配[J]. 动物学报, 53(4): 589-599. |
黄勇, 权锐昌, 任国鹏, 等. 2008. 朱建国西藏米拉卡黑白仰鼻猴的栖息地变化[J]. 动物学研究, 29(6): 653-660. |
蒋志刚. 2004. 动物行为原理与物种保护方法[M]. 北京: 科学出版社.
|
李明晶, 马建章. 1989. 麻阳河自然保护区黑叶猴生态及其数量的初步调查[J]. 野生动物, (4): 13-15. |
李明晶. 1994. 贵州野生动物资源[M]. 北京: 中国林业出版社: 28-30.
|
李言, 张明海, 蒋志刚. 2008. 基于生境可获得性的完达山地区马鹿(Cervus elaphus xanthopygus)冬季生境选择[J]. 生态学报, 28(10): 4619-4628. DOI:10.3321/j.issn:1000-0933.2008.10.001 |
李友邦, 黄乘明, 黄中豪, 等. 2008. 扶绥黑叶猴食物多样性的初步研究[J]. 广西师范大学学报:自然科学版, 26(1): 96-99. |
刘璐. 2009. 宽阔水国家级自然保护区黑叶猴生境利用的初步观察[M]. 贵阳: 贵州师范大学生命科学学院硕士学位论文: 1-29.
|
刘万福, 韦振逸.1995.广西灵长类资源与保护//夏武平, 张荣祖.灵长类研究与保护.北京:中国林业出版社, 123—132.
|
刘瑛, 刘广超, 陈建伟. 2009. 灾后川金丝猴适宜栖息地恢复指标的研究[J]. 西部林业科学, 38(2): 33-40. |
黎大勇, 彭正松, 任宝平, 等. 2006. 塔城滇金丝猴初秋对生境的选择性[J]. 西华师范大学学报:自然科学版, 27(3): 233-238. |
罗杨, 陈正仁, 汪双喜. 2000. 贵州麻阳河地区黑叶猴的食性观察[J]. 动物学杂志, 35(3): 44-45. |
罗杨, 张明海, 马建章, 等. 2005. 贵州黑叶猴日活动时间的分配[J]. 兽类学报, 25(2): 156-162. |
罗杨, 张明海, 马建章, 等. 2007. 干扰生境下黑叶猴(Trachypithecus francoisi francoisi)日活动时间的分配[J]. 生态学报, 27(5): 1715-1722. |
罗振华, 刘丙万, 刘松涛. 2008. 内蒙古达赉湖地区蒙原羚的春季生境选择[J]. 兽类学报, 28(4): 342-352. |
马建伦, 谢章桂. 2007. 金佛山国家级自然保护区综合考察初报[J]. 重庆林业科技, (2): 25-30. |
马建章, 邹红菲, 贾竞波. 2004. 野生动物管理学[M]. 2版. 哈尔滨: 东北林业大学出版社: 43-62.
|
尚玉昌. 1998. 行为生态学[M]. 北京: 北京大学出版社: 240-252.
|
孙儒泳. 2001. 动物生态学原理[M]. 3版. 北京: 北京师范大学出版社: 271-272.
|
苏化龙, 林英华, 马强, 等. 2002. 重庆市武隆县和彭水县交界处白颊黑叶猴种群初步调查[J]. 兽类学报, 22(3): 169-178. |
唐洪通, 张含藻. 1998. 白颊黑叶猴的保护与利用[J]. 四川林业科技, (2): 49-51. |
王应祥, 蒋学龙, 冯庆. 1999. 中国叶猴类的分类、现状与保护[J]. 动物学研究, 20(4): 306-315. |
魏辅文, 冯祚建, 王祖望. 1998. 野生动物对生境选择的研究概况[J]. 动物学杂志, 33(4): 48-52. |
吴名川, 韦振逸, 何农林. 1987. 黑叶猴在广西的分布及生态[J]. .野生动物, (4): 12-19. |
谢东明, 路纪琪, 吕九全. 2009. 太行山猕猴的冬季生境选择[J]. 兽类学报, 29(3): 252-258. |
颜忠诚, 陈永林. 1998. 动物的生境选择[J]. 生态学杂志, 17(2): 43-49. |
叶智彰. 1993. 叶猴生物学[M]. 昆明: 云南科技出版社: 32-42.
|
张含藻, 刘正宇, 胡周强, 等. 1992. 金佛山自然保护区首次发现白颊黑叶猴[J]. 四川动物, 11(4): 30. |
张含藻, 刘正宇, 胡周强, 等. 1994. 白颊黑叶猴资源及生态调查[J]. 中药材, 17(4): 11-13. |
张洪海, 马建章. 1999. 紫貂冬季生境的偏好[J]. 动物学研究, 20(5): 355-359. |
张鹏, 李保国, 渡边邦夫. 2006. 秦岭川金丝猴冬季和春季在自然栖息地的空间利用[J]. 动物学报, 52(3): 429-436. |
张强, 兰盛军, 胡大明, 等. 2008. 四川白水河国家级自然保护区野外藏酋猴活动初步调查[J]. 四川动物, 27(1): 131-134. |
张文彤. 2002. SPSS11统计分析教程(高级篇)[M]. 北京: 希望电子出版社: 91-109.
|
周岐海, 黄乘明, 方艳. 2001. 黑叶猴春夏季活动时间的初步研究[J]. 广西师范大学学报:自然科学版, l9(4): 80-83. |
周岐海. 2004. 弄岗黑叶猴对喀斯特石山生境的行为适应[M]. 北京: 中国科学院动物研究所博士学位论文: 66-79.
|
周岐海, 黄乘明, 李友邦. 2007a. 黑叶猴活动时间季节性变化[J]. 动物学杂志, 42(1): 67-73. |
周岐海, 蔡湘文, 黄乘明, 等. 2007b. 黑叶猴在喀斯特石山生境的觅食活动[J]. 兽类学报, 27(3): 243-248. |
周岐海, 黄中豪, 韦华, 等. 2009. 同域分布黑叶猴和熊猴的活动时间分配比较[J]. 兽类学报, 29(1): 1-6. |
周汝良, 杜勇, 杨庆仙, 等. 2008. 滇金丝猴栖息地的空间格局分析[J]. 云南地理环境研究, 20(3): 1-5. |
Ables E D. 1969. Home range studies of red foxes (Vulpes vulpes)[J]. J Mamm, 50(1): 108-120. DOI:10.2307/1378635 |
Baily J A. 1984. Principles of Wildlife Management[M]. New Yok: John Wiley and Sons, Inc.
|
Block W M, Brennan L A. 1993. The habitat concept in ornithology: theory and applications[J]. Cur Orni, 11: 35-91. |
Boonratana R. 2000. Ranging behavior of proboscis monkey(Nasalis larvatus) in the lower Kinabalangan, Northern Borneo[J]. Int J Primatol, 21: 497-518. DOI:10.1023/A:1005496004129 |
Boyce M S, McDonald L L. 1999. Relation populations to habitats using resource selection functions[J]. Trends in Ecology and Evolution, 14: 268-272. DOI:10.1016/S0169-5347(99)01593-1 |
Chapman C A, Chapman L J, MeLaughlin R L. 1989. Multiple central place foraging by spider monkeys: travel consequences of using many sleeping sites[J]. Oecologia, 79: 506-511. DOI:10.1007/BF00378668 |
Cowlishaw G. 1997. Trade-offs between foraging and predation risk determine habitat use in a desert baboon population[J]. Anim Behav, 53: 667-686. DOI:10.1006/anbe.1996.0298 |
Cyril C G, Ding Wei. 2006. An observation of Francois' Langur using caves at Mayanghe National Nature Reserve, Guizhou, China[J]. Zoological Research, 27(5): 558-560. |
Enstam K L, Isbell L A. 2004. Microhabitat preference and vertical use of space by patas monkeys(Erythrocebus paras)in relation to predation risk and habitat structure[J]. Folia Primatol, 75: 70-84. DOI:10.1159/000076265 |
Harrison M J S. 1983. Patterns of range use by the green monkey Cercopithecus sabaeu at Mt[J]. Assirik, Senega1. Folia Primatol, 41: 157-179. DOI:10.1159/000156129 |
Hosmer Jr D W, Lemeshow S. 1989. Applied logistic Regression[M]. New York: John Wiley & Sons.
|
Huang Chengming, Li Youbang, Zhou Qihai. 2003. Activity patterns and their occurrence in day range of Francois Langur (Trachypithecus francoisi) in Fusui Reserve, Guangxi[J]. Journal of Guangxi Normal University, 2l(4): 78-82. |
Kaplin B A. 2001. Ranging behavior of two species of guenons(Cercopithecus lhoesti and C. mitis doggetti) in the Nyungwe Forest Reserve[J]. Rwanda. Int J Primatol, 22: 521-548. DOI:10.1023/A:1010716001014 |
Lechowicz M J. 1982. The sampling characteristics of selectivity indices[J]. Ecology, 52(1): 22-30. |
Lennon J J. 1999. Resource selection functions: taking space seriously?[J]. Trends in Ecology and Evolution, 14: 399-400. |
Lima S L, Dill L M. 1990. Behavioural decisions under the risk of predation:a review and prospectus[J]. Can J Zool, 68: 619-640. DOI:10.1139/z90-092 |
Manly B F J, McDonald L L, Thomas D L. 1993. Resource Selection by Animals: statistical Design and Analysis for Field Studies[M]. London: Chapman & Hall.
|
McGrew W C, McKee J K, Tutin C E G. 2003. Primatesin caves:two new reports of Papio spp[J]. J Hum Evol, 44: 521-526. DOI:10.1016/S0047-2484(03)00042-3 |
McLoughlin P D, Case R L, Gau R J, et al. 2002. Hierarchical habitat selection by barren-ground grizzly bears in the central Canadian Arctic[J]. Oecologia, 132(1): 102-108. DOI:10.1007/s00442-002-0941-5 |
Olupot W, Chapman C, Waser P M, et al. 1997. Mangabey(Cercocebus albigena) ranging patterns in relation to fruit availability and the risk of parasite infection in Kibale National Park, Uganda[J]. Am J Primato1, 43: 65-78. DOI:10.1002/(ISSN)1098-2345 |
Zhou Qihai, Wei Fuwen, Li Ming. 2006. Diet and Food Choice of Trachypithecus francoisi in the Nonggang Nature Reserve, China[J]. International Journal of Primatology, 27(5): 1441-1460. DOI:10.1007/s10764-006-9082-8 |
Su Y J, Ren R M, Yan K H, et al. 1998. Preliminary survey of the home range and ranging behavior of golden monkeys(Rhinopithecus roxellena) in Shennongjia National Natural Reserve, Hubei, China//Jablonski N G The Natural History of the Doucs and Snub nosed Monkeys[M]. Singapore: World Scientific Publishing: 255-268.
|
Suzuki A. 1965. An ecological study of wild Japanese monkeys in snowy areas focused on their food habits[J]. Primates, 6(1): 31-72. DOI:10.1007/BF01794458 |
Wada K. 1964. Some observations on the life of monkeys in a snowy district of Japan[J]. Physiology and Ecology, 12: 151-174. |
Wu Gang, Wang Hong chang, Fu Hai wei, et al. 2004. Habitat selection of Guizhou golden monkey(Phinopithecus roxellanae brelichi)in Fanjing Mountain Biosphere Reserve[J]. China. Journal of Forestry Research, 15(3): 192-202. DOI:10.1007/BF02911023 |