 2011, Vol. 47
2011, Vol. 47文章信息
- 张克荣, 刘应迪, 朱晓文, 张全发, 谭淑端
- Zhang Kerong, Liu Yingdi, Zhu Xiaowen, Zhang Quanfa, Tan Shuduan
- 长沙岳麓山马尾松林的群落类型划分及物种多样性分析
- Community Types and Species Diversity of Pinus massoniana Forests of Yuelu Mountain, Changsha
- 林业科学, 2011, 47(4): 86-94.
- Scientia Silvae Sinicae, 2011, 47(4): 86-94.
-
文章历史
- 收稿日期:2009-06-29
- 修回日期:2010-12-21
-
作者相关文章



2. 中国科学院武汉植物园水生植物与流域生态重点实验室 武汉 430074;
3. 中国科学院研究生院 北京 100049
2. Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, The Chinese Academy of Sciences Wuhan 430074;
3. Graduate University of the Chinese Academy of Sciences Beijing 100049
随着全球城市化水平的迅速提高, 环境问题日益加剧, 城市森林的地位与作用、保护与经营等越来越受到重视(McPherson et al., 1997; 罗慧君等, 2004; 彭羽等, 2007)。研究城市森林的群落结构及物种多样性, 可为人工群落树种选择及配置、城市森林空间格局的优化、城市森林景观异质性的改善、城市植被的科学管理等提供理论依据(童丽丽等, 2007; 张克荣等, 2009a)。
植物群落物种多样性关系到生态系统生产力、稳定性、可入侵性等(Brown et al., 2003; Balvanera et al., 2006; Cardinale et al., 2006; 郑景明等, 2006; 郭轶瑞等, 2007)。目前对于退化生态系统的恢复和重建, 越来越强调生物多样性和生态功能的恢复(Lamb et al., 2005)。松科的一些树种由于其较强的适应能力, 在全世界范围内被广泛用于生态恢复和植树造林。但很多研究表明, 一些松树人工林造成土壤酸化、养分耗竭、生物地球化学循环紊乱、病虫害暴发、生物多样性下降等问题(Liu et al., 1998; Scholes et al., 1998; 孟庆繁, 2006; 王希华等, 2001), 因此认为松树人工林不利于生物多样性保护。另一方面, 有很多研究者却得出截然不同的观点, 他们认为松树人工林在生物多样性保护中起着非常重要的作用, 能够为乡土物种提供庇护所或合适的替代生境, 能够促进演替中后期树种的建立, 其林下本土物种与天然林中一样丰富(Ashton et al., 1997; Oberhauser, 1997; Brockerhoff et al., 2003; Czerepko, 2004; 孟庆繁, 2006; Langer et al., 2008)。
马尾松(Pinus massoniana)具有耐干旱、耐瘠薄、适应能力强等特点, 被广泛用于我国荒山造林, 其造林面积仅次于杉木(Cunninghamia lanceolata)人工林, 位居第2位(王希华等, 2001; 项文化等, 2002)。虽然马尾松人工林对于退化生态系统恢复起到积极作用, 但大面积马尾松纯林容易引起病虫害暴发流行(王希华等, 2001), 从而使生态系统服务功能下降, 并且由于杀虫剂的大量使用, 造成新的环境问题(田晔林等, 2002)。近年来, 已有许多学者对马尾松林进行过研究, 涉及种群动态(杨心兵等, 2001)、群落结构(王献溥等, 2002; 童群益等, 2004)、演替机制(丁圣彦等, 1998; 王希华等, 2001)、生物量和生产力(丁贵杰等, 2001)、养分循环特征(项文化等, 2002)、病虫害暴发原因(罗永松, 1997)、经营管理及改造技术等多方面(童群益等, 2004; 康冰等, 2005; 林德喜等, 2005), 马尾松林群落类型的划分也有报道, 例如童群益等(2004)将浙江马尾松林划分为8种类型, 王献溥等(2002)将广西马尾松林分为76个群丛, 但这些分类都有一定主观性, 对马尾松林进行数量分类和排序鲜见报道, 尤其是对中亚热城市森林中马尾松群落了解不够。植物群落的分类和排序是进行植物群落多样性研究的基础(丁圣彦等, 1998), 了解城市森林中马尾松林的群落类型、群落结构及物种多样性特征, 对于合理经营管理马尾松林、维持城市生态系统健康尤为重要。
岳麓山国家级风景名胜拥有长沙市区内仅存的一片自然植被和保存较完好的城市森林, 对于改善和维持长沙市的城市生态环境具有十分重要的作用。马尾松是岳麓山乔木层的优势种之一(张克荣等, 2009a), 马尾松林的群落学特征强烈影响岳麓山生态系统功能的发挥。本文以岳麓山的马尾松林为研究对象, 运用TWINSPAN, flexible beta clustering(可变聚合法), DCA, MDS(无度量多维标定排序, non-metric multidimensional scaling)马尾松林进行分类和排序, 并分析群落结构及物种多样性, 旨在为科学管理马尾松林、构建合理人工群落提供指导, 并对以上4种多元分析方法的应用效果进行比较。
1 研究地区和研究方法 1.1 研究地区的自然概况岳麓山位于112°44′—112°48′ E, 28°20′—28°27′ N, 南北长约4 km, 东西宽约1.5~2 km, 山体面积6 km2, 主峰海拔300.8 m(田晔林等, 2002), 母岩为石英砂岩、页岩、砂岩, 主要土壤类型为红壤、山地黄红壤(张明, 1997), 表土多为腐殖质层。长沙属亚热带湿润气候区, 年平均降雨1 200~1 400 mm(田晔林等, 2002), 4—7月降水最多, 为全国多雨区之一。夏季高温多雨, 春夏之际阴湿多雨, 秋末冬初雨量较少, 冬季气候冷湿, 年平均相对湿度为80%, 年平均气温为17.2 ℃, 年平均最低气温4.7 ℃(1月), 年平均最高气温29.5 ℃(7, 8月), 年较差24.8 ℃(黄宗胜等, 2005)。
岳麓山地处中亚热带常绿阔叶林地带, 植被主要由天然群落构成。自然植物群落经历几次较大的人为干扰:1951—1955年, 人工栽培大量马尾松、枫香(Liquidambar formosana)(彭映辉等, 1992); 1965—1966年由湖南林校部分学生进行一次较大规模人工筛选, 淘汰部分植物(张明, 1997)。1975年建成公园后, 由于禁止开山采石、葬坟修墓、违章搭建、伐木毁林、带入火种、猎采动植物, 干扰程度降低, 植被得到较好的保护。
1.2 调查取样2006年, 在对岳麓山全面踏查的基础上, 确定一条围绕整个山体的横向路线, 样地间隔约500 m, 再在每个与取样位置垂直的路线上取样2~3个, 具体数目根据从山脚至山脊或山顶的相对高度来确定(海拔间隔约50 m左右), 但确保每个坡向的高、中、低海拔至少有1个样地。共调查43个样地, 乔木层样方面积为20 m×20 m, 样地总面积占岳麓山山体面积的0.29%。用GPS记录样地位置、海拔等, 将高度>3 m, 且胸径≥2.5 cm的植物记为乔木层, 对它们进行每木调查, 记录种名、胸径、高度、枝下高、冠幅等。在每个样地内随机取2个6 m×6 m灌木层样方, 共86个; 另外随机取4个2 m×2 m草本层样方, 共172个。灌木层和草本层样方记录种类、数量、高度、盖度等, 层间植物归人灌木层和草本层。选取其中28个有马尾松的样地进行分析, 样地概况列于表 1。
|
|
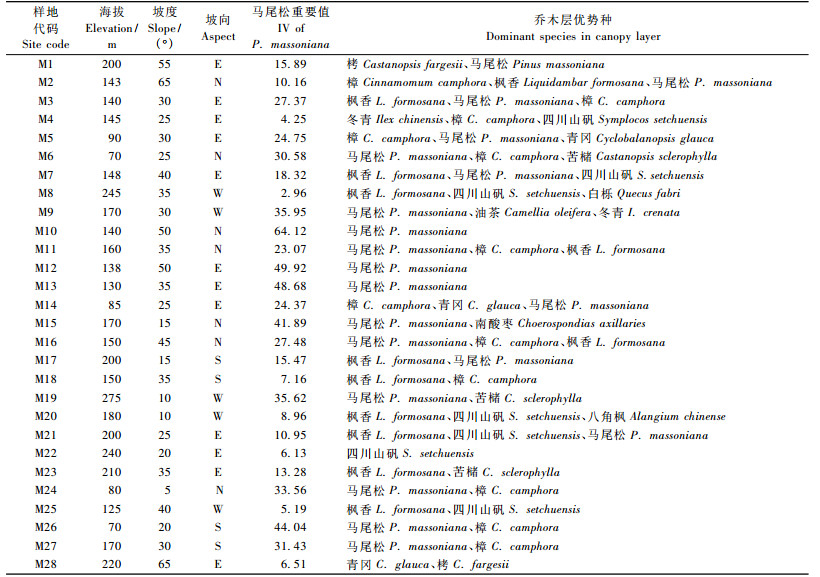
乔木层物种相对重要值=(相对密度+相对优势度+相对频度)×100/3, 灌木层相对重要值=(相对密度+相对盖度+相对频度)×100/3, 草本层物种相对重要值=(相对盖度+相对频度)×100/2。其中乔木层的相对优势度使用胸径的数据计算得出。
1.3.2 群落分类与排序样地与物种重要值组成二维数据表作为分类和排序的原始矩阵, TWINSPAN, flexible beta clustering, DCA, NMDS在PC-ORD for Windows (version 4.0)上完成。运行程序时, TWINSPAN中将pseudospecies cut levels设置0, 2, 5, 10, 20, 30, 40, 50, flexible beta clustering中取beta = -0.25, DCA计算中, 把down weight rare species选中, 其他均采用默认值, 具体运算过程见相关文献(张金屯, 2004)。
1.3.3 群落的物种多样性指数本文选用以下几个使用较多的指数分析物种多样性(张金屯, 2004):丰富度Patrick指数S; 优势度采用Simpson指数:D = 1-Σ Pi2; 多样性采用Shannon-Wiener指数:H =-Σ PilnPi; 均匀度采用Pielou指数:E = H/lnS =-ΣPilnPi/lnS。以上公式中, S为样方中的物种数目, Pi为种i的重要值。
2 结果与分析 2.1 群落类型的划分TWINSPAN将28个样地划分为9类(图 1)。DCA三轴的特征值分别为X1=0.230 5, X2=0.174 9, X3=0.121 7, 三轴累积贡献率为25.8%, DCA第一轴主要反映海拔的变化(r = 0.695, P < 0.001), 其他轴反映的环境意义不明显。TWINSPAN划分的群落类型大部分能够在DCA排序图(图 2)上反映出来, 但对于M10, M16, M21, M23样地的处理存在较大的差异, TWINSPAN将M10与M2, M10, M26聚为一类, 而根据DCA排序图(图 2), 倾向与将M4, M6, M7, M10划分为同一类群落, 而将M16, M2, M14, M26分为同一群落。

|
图 1 岳麓山28个样地TWINSPAN分类 Figure 1 TWINSPAN classification of 28 plots in Yuelu Mountain |

|
图 2 岳麓山28个样地DCA二维散点图(X1与X2) Figure 2 Two-dimensional scatter plot of DCA ordination for 28 plots in Yuelu Mountain |
图 3, 4分别是flexible beta clustering和NMDS的结果。当保留70%左右信息时, flexible beta clustering与NMDS的结果十分一致, flexible beta clustering将28个样地分成13种类型(图 3), 每一种类型在NMDS排序图上都能明显区分开(图 4中实线图形所示)。

|
图 3 岳麓山28个样地的flexible beta clustering聚类 Figure 3 Flexible beta clustering for 28 plots in Yuelu Mountain |

|
图 4 岳麓山28个样地的NMDS排序 Figure 4 NMDS ordination for 28 plots in Yuelu Mountain |
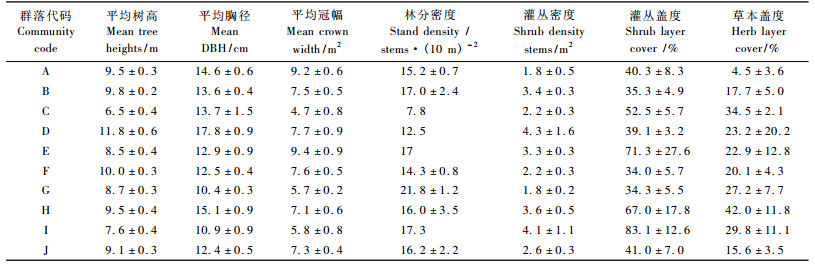
分析各样地优势种特征(表 1), M12为马尾松占绝对优势的纯林, 因此将同样是该种植被类型的M10, M13归为同一种类型(图 4)。M27, M3, M16, M26, M11样地的优势种为马尾松、枫香、樟, 将这5个样地并为一类, 同样将以樟、马尾松为优势种的M5与M6, M24, M2, M14, M4并为一类。这种合并在NMDS排序图得到反映(图 4), 并且NMDS排序图上各群落类型的区分更加明显。最后得到10种群落类型(表 2, 图 4), 根据《中国植被》分类的原则划分为暖性常绿针叶林、亚热带山地落叶阔叶林、亚热带常绿阔叶林和温性针叶阔叶混交林4种植被型, 10个群系(表 2, A~J), 其中7群系中马尾松为优势种。
|
|

岳麓山马尾松存在于10种群系中(表 2), 经过50年的群落演替, 岳麓山仍然存在较大面积的马尾松纯林(10.7%样地为马尾松纯林), 但更多的是马尾松与樟、枫香、苦槠、四川山矾、南酸枣、油茶等阔叶树种形成混交林。
2.2 群落结构特征各群落结构的基本特征列于表 3。马尾松纯林主要由人工种植而来, 马尾松为乔木层的建群种, 有少量青冈、樟、冬青、白栎进入乔木层。平均林分密度为15.2株·(10 m)-2。群落结构较简单, 林下灌木以稀疏的檵木(Loropetalum chinensis)、乌饭树(Vaccinium bracteatum)为主, 草本植物极为稀疏(表 3), 以阔鳞鳞毛蕨(Dyropteris championi)为优势种。藤本植物有羊角藤(Morinda umbellate)、南蛇藤(Celastrus articulatus)、菝葜(Smilax china)、土茯苓(Smilax globra)等。阔叶树种樟、青冈、石栎(Lithocarpus glaber)、栲、苦槠等的幼树和幼苗也出现在林下, 但数量不多。
|
|
马尾松+樟+枫香林乔木层成分较复杂, 除3种优势乔木树种外, 尚有白栎、冬青、南酸枣等, 并且格药柃(Eurya muricata)、八角枫(Alangium chinense)、四川山矾等小乔木形成乔木亚层, 样地中平均胸径为12.3~15.6 cm的下层乔木丰富; 以油茶、白花苦灯笼(Tarenna mollissima)、黄栀子(Gardenia jasminoides)为优势灌木, 樟幼苗较多; 草本植物以阔鳞鳞毛蕨、淡竹叶(Lophatherum gracile)等为主。藤本植物有鸡矢藤(Paederia scandens)、南蛇藤、菝葜、土茯苓、海金沙(Lygodium japonicum)等。
马尾松+苦槠群落林分密度小为7.8株·(10 m)-2, 林下植被发育良好, 种类非常丰富, 以毛叶木姜子(Litsea euosma)、菝葜、淡竹叶等为优势种, 苦槠、栲、樟、青冈等阔叶树种幼树很多, 体现向地带性植被亚热带常绿阔叶林演替的趋势。
马尾松+南酸枣林平均胸径较大(17.8 cm), 乔木层混生有白栎、枫香、樟等, 灌丛密度大(4.3株·m-2), 白花苦灯笼、大青叶(Clerodendrum cyrtophyllum)、山莓(Rosa corchoriferus)为优势灌木, 愉悦蓼(Polygonum jucundum)、淡竹叶为优势草本, 林下樟树幼苗丰富。
马尾松+油茶林是由马尾松侵入废弃的油茶林发展而来, 乔木层中尚有冬青、樟、枫香等。林下灌木油茶、花竹(Phyllostachys nidularia)为优势, 芒(Miscanthus sinensis)、蕨(Pteridium aquilinum var.latiusculum)为优势草本, 灌木层盖度大(71.3%), 藤本植物有鸡血藤(Milletia reticulata)、菝葜、金樱子(Rosa laevigata)等。
马尾松+樟林是一种广泛分布于岳麓山的群落, 乔木层中除樟、枫香外, 冬青、苦槠、青冈、白栎、黄瑞木(Adinandra millettii)为主要主要伴生种。林下灌木层以白花苦灯笼、山矾(Symplocos sumumtia)、大青叶、花竹等为优势种。樟、青冈、石栎幼苗幼树丰富。狗脊(Woodwardia japonica)、阔鳞鳞毛蕨、淡竹叶为优势草本。
枫香+马尾松+四川山矾群落林分密度大为21.8株·(10 m)-2, 且分层明显, 枫香、马尾松为第1乔木层优势种, 四川山矾为第2乔木层优势种, 并伴生有八角枫、格药柃、冬青等。大青叶、毛叶木姜子、四川山矾(幼树)等为优势灌木, 狗脊、阔鳞鳞毛蕨、淡竹叶为草本层优势种。藤本植物有鸡血藤、菝葜等。
2.3 马尾松群落的物种多样性岳麓山马尾松林(马尾松为优势种或共优种的7种群落, 即马尾松纯林和针阔混交林)21个样地共记录到维管植物101种, 其中乔木35种, 灌木71种(包括乔木幼树28种), 草本23种。各群落的多样性指数列于表 4。
|
|
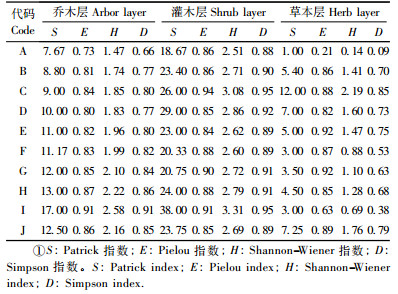
四川山矾林乔木层丰富度指数最高, 其次是栲+青冈林、枫香林。几种针阔混交林中, 乔木层物种丰富度指数大小为:枫香+马尾松+四川山矾林>马尾松+樟林>马尾松+油茶林>马尾松+南酸枣林>马尾松+苦槠林>马尾松+樟+枫香林; 灌木层物种丰富度大小为马尾松+南酸枣林>马尾松+苦槠林>马尾松+樟+枫香林>马尾松+油茶林>枫香+马尾松+四川山矾林>马尾松+樟林; 草本层物种丰富度大小为:马尾松+苦槠林>马尾松+南酸枣林>马尾松+樟+枫香林>马尾松+油茶林>枫香+马尾松+四川山矾林>马尾松+樟林。马尾松纯林的乔木层、灌木层、草本层丰富度指数均为最低值。
各样地均匀度指数变化不大, 但马尾松林乔木层和草本层均匀度指数最低。四川山矾林乔木层多样性指数最高。对于灌木层而言, 马尾松+苦槠林均匀度最高。
马尾松纯林乔木层、灌木层、草本层多样性指数均为最低值。四川山矾林、栲+青冈林、枫香林的乔木层中多样性指数均高于针阔混交林, 四川山矾林灌木层中多样性指数最高, 其次是马尾松+苦槠林。
马尾松+苦槠林灌木层、草本层中优势度指数最大。对于乔木层而言, 四川山矾林优势度指数最大, 其次是栲+青冈林、枫香+马尾松+四川山矾, 乔灌草3层的优势度最小值均出现在马尾松林中。
2.4 马尾松重要值与物种多样性的关系将28个样地中马尾松的重要值与各样地物种多样性指数进行相关分析, 结果显示, 样地中马尾松的重要值与乔木层的物种丰富度指数、均匀度指数、多样性指数、优势度指数均存在极显著的负相关, 相关系数分别为-0.618 (P=0.000), -0.774 (P =0.000), -0.766 (P =0.000), -0.839 (P =0.000), 而与灌木层的物种丰富度指数、均匀度指数、多样性指数、优势度指数均不存在相关关系(P >0.05), 但与草本层的均匀度指数、优势度指数存在负相关, 相关系数分别为-0.525 (P =0.004), -0.467 (P =0.012), 与草本层的丰富度指数、多样性指数不存在相关关系(P >0.05)。
这一分析表明, 马尾松对乔木层物种多样性具有强烈的主导作用, 对于灌木层和草本层而言, 这一作用则不那么显著。
3 讨论TWINSPAN和DCA被认为是目前较为完善和应用效果较理想的植物群落分类和排序的方法, 在植被研究中占主导地位(胡理乐等, 2005; 曹伟等, 2007), 但都存在缺点(张金屯, 2004)。对于岳麓山马尾松林的应用结果表明, 相对于flexible beta clustering和NMDS, TWINSPAN和DCA的结果并不理想。主要表现在TWINSPAN和DCA的结果存在较大差异, 尤其是TWINSPAN对于马尾松占绝对优势M10样地的处理(表 1, 图 1), 可以认为是错分样地, flexible beta clustering, NMDS, DCA的结果都证明这点(图 2, 3, 4)。DCA三轴累积贡献率很低, 只有25.8%。相比之下, flexible beta clustering和NMDS的结果十分一致, 尤其是在保留70%信息时, 分类与排序的结果完全一致(图 3, 4)。综合考虑样地的生境与物种组成特征, 最终将28个样地分成10组。10组群落在NMDS排序图上具有明显的边界, 代表 10种具有不同优势种的群系, 划分结果具有明确的生态学意义, 是客观合理的(图 4, 表 1, 2)。胡理乐等(2005)对辽东山区天然次生林的研究也表明TWINSPAN会产生边界样地、错分样地。虽然flexible beta clustering和NMDS在群落数量分类中使用频率较低(张金屯, 2004), 但本研究的结果表明, flexible beta clustering和NMDS相结合要优于TWINSPAN和DCA, 是比较理想的群落数量分类方法。
本试验分类和排序的结果表明, 岳麓山马尾松除组成较大面积的纯林外, 还与其他本土阔叶树种组成6种针阔混交林(表 2)。岳麓山针阔混交林的来源和形成途径主要有2种:一是本土阔叶树种侵入马尾松人工林中形成共优种, 二是马尾松通过自然扩散, 进入本土树种形成的天然次生林中。2种途径均为受损生态系统(被破坏的天然林)在干扰程度降低后, 经过30~50年演替形成, 一定程度上反映岳麓山马尾松林的演替趋势:针叶林→针阔混交林→常绿阔叶林(张克荣, 2009a)。根据自我设计理论, 这类生态系统的组分和结构是退化生态系统根据环境条件合理组织所形成的(任海等, 2004)。因此本研究所揭示的岳麓山马尾松林的群落组分、结构及动态特征, 对于本地区退化生态系统恢复、人工群落配制模式、城市绿地规划与管理等具有指导意义。
本研究的结果表明, 马尾松纯林物种多样性极低, 不仅是乔木层, 灌木层和草本层丰富度、多样性均为最低值。这种情况与世界上其他一些对于松树人工林林下本土植物多样性的研究不同, 如新西兰的辐射松(Pinus radiata)(Brockerhoff et al., 2003; Langer et al., 2008)、斯里兰卡的加勒比海松(Pinus caribaea)(Ashton et al., 1997), 波兰的欧洲赤松(Pinus sylvestris)(Czerepko, 2004)、泰国的Pinus kesiya (Oberhauser, 1997), 这些研究表明松树人工能够形成本土物种极为丰富的林下层。马尾松作为一种亚热带地区的本土先锋种, 为什么会表现出这种差别?这种差异是否与造林密度、立地条件、管理方式等有关?值得深入研究。
马尾松的重要值与乔木层的各物种多样性指数、草本层的均匀度指数、优势度指数存在负相关, 而与灌木层的物种多样性不存在相关关系, 暗示马尾松对乔木层中的物种多样性具有强烈的主导作用, 而对于灌木层而言, 这一作用则不那么明显。当然马尾松也可能对林下草本的均匀度、优势度产生影响, 马尾松通过决定群落自身的结构特征来影响群落的物种多样性。样地调查时观察到, 马尾松纯林下草本十分稀少, 3个样地12个草本样方中, 只调查到阔鳞鳞毛蕨、栗褐苔草(Carex brunnea)2种植物, 草本层平均盖度只有4.5%, 但样地中马尾松林大树却很多(马尾松主要为1951—1955年人工种植, 1975年开辟为公园后, 人为干扰较少), 且长势良好, 枯枝落叶层、腐殖质层深厚。虽然很多学者已经注意到马尾松林物种多样性较低(罗永松, 1997), 但对其具体原因和机制却了解不够。考虑到群落年龄与样地特征, 本研究所显示的马尾松纯林草本层物种多样性极低的现象, 很难完全从马尾松林处于次生演替先锋阶段(丁圣彦等, 1998; 王希华等, 2001)、地力衰退等原因(王巧珍, 2003)得到解释。近年来许多研究表明, 枯枝落叶层通过物理阻断、化感作用等对森林的天然更新产生障碍(王贺新, 2008)。在样地调查中注意到, 马尾松纯林下积累大量凋落物, 这主要与马尾松的枯叶和枯枝的周转时间普遍大于阔叶树有关(林德喜等, 2005)。马尾松纯林草本层极低的物种多样性是否与凋落物有关值得进一步研究。如果马尾松纯林下凋落物对于物种多样性的恢复存在一定程度的抑制作用, 那么根据“强调生物多样性和生态功能恢复的理念”(Lamb, 2005), 对我国现有的大面积马尾松人工林的管理将提出新的课题, 当然这有待更深入的研究。
罗永松(1997)对赣南马尾松的研究表明, 马尾松林分中灌木层物种多样性与松毛虫暴发存在负相关, 并根据模型, 认为要保持马尾松毛虫不成灾, 灌木层物种多样性指数应大于或等于2.67。本研究结果显示(表 4), 岳麓山马尾松纯林灌木层多样性指数(H)只有2.51, 虽然不同的地域之间存在差异, 但是本研究所揭示的马尾松林低物种多样性的特征, 至少为岳麓山马尾松毛虫暴发(田晔林等, 2002)的原因提供一种可能的解释。相比之下, 针阔混交林, 尤其是马尾松+苦槠林, 则显示出较高的物种多样性, 这主要是由于马尾松+苦槠林灌木层具有较高的物种多样性(H=3.08)、草本层具有较高的物种丰富度(S=12), 表明马尾松+苦槠林下植被发育良好, 这与王巧珍(2003)的研究结果类似。另外, 考虑到苦槠是岳麓山地带性植被亚热带常绿阔叶林的代表树种(彭映辉等, 1992), 马尾松+苦槠林可视为岳麓山马尾松林向地带性植被恢复过程中的一个过渡阶段。近年来已有许多学者提出将马尾松林向针阔混交林、常绿阔叶林改造, 以提高生态系统的服务功能(王希华等, 2001; 王巧珍, 2003; 林德喜等, 2005)。可以推测, 如果将岳麓山马尾松纯林改造称为针阔混交林, 将可提高其物种多样性, 并促进其向常绿阔叶林演替, 从而提高生态系统的稳定性和生态服务功能(王希华等, 2001; 康冰等, 2005; 林德喜等, 2005)。
长沙是我国“两型社会”试验区之一, 处在城市化水平的迅速提高时期。城市建设、交通发展的加快必定会造成部分生态系统退化, 如新的边坡、裸地的形成。在“两型社会”建设过程中, 退化生态系统恢复、城市森林构建与科学管理等问题必须引起高度重视, 才能实现“环境友好型社会”。虽然马尾松是中亚热带植被演替过程中的消退种(丁圣彦等, 1998; 张克荣等, 2009b), 且马尾松林(尤其是纯林)存在物种多样性低、易暴发病虫害等弊病, 但毕竟生态系统恢复首要的是阻止进一步退化(如水土流失等)(Lugo, 1997)。因此, 马尾松作为本气候带最主要的生态恢复本土先锋种和驱动种之一(张克荣等, 2009a), 仍将发挥至关重要的作用, 尤其是在一些退化较严重的生态系统中, 马尾松的作用不容忽视, 但必须构建科学合理的人工群落。
蔡聪. 1995. 马尾松阔叶树混交林物种多样性的初步研究[J]. 中南林学院学报, 15(2): 195-200. |
曹伟, 李岩, 王树良, 等. 2007. 东北阔叶红松林群落类型划分及物种多样性[J]. 应用生态学报, 18(11): 2406-2411. |
丁贵杰, 王鹏程. 2001. 马尾松人工林生物量及生产力变化规律研究:Ⅱ.不同林龄生物量及生产力[J]. 林业科学研究, 15(1): 54-60. |
丁圣彦, 宋永昌. 1998. 常绿阔叶林演替过程中马尾松消退的原因[J]. 植物学报, 40(8): 755-760. |
高贤明, 陈灵芝. 1998. 北京山区辽东栎(Quercus liaotunensis)群落多样性的研究[J]. 植物生态学报, 22(1): 23-32. |
郭轶瑞, 赵哈林, 赵学勇, 等. 2007. 科尔沁沙质草地物种多样性与生产力的关系[J]. 干旱区研究, 24(2): 199-203. |
胡理乐, 毛志宏, 朱教君, 等. 2005. 辽东山区天然次生林的数量分类[J]. 生态学报, 25(11): 2848-2854. DOI:10.3321/j.issn:1000-0933.2005.11.008 |
黄宗胜, 彭重华, 王建兵. 2005. 岳麓山风景名胜区枫香群落研究[J]. 湖南林业科技, 32(3): 24-26. |
康冰, 刘世荣, 史作民, 等. 2005. 南亚热带人工马尾松林下植物组成特征及主要木本种群生态位研究[J]. 应用生态学报, 16(9): 1786-1790. |
林德喜, 樊后保. 2005. 马尾松林下补植阔叶树后森林凋落物量、养分含量及周转时间的变化[J]. 林业科学, 41(6): 7-15. DOI:10.11707/j.1001-7488.20050602 |
罗慧君, 包志毅. 2004. 欧洲城市林业概述[J]. 世界林业研究, 17(3): 44-47. |
罗永松. 1997. 灌木层物种多样性与松毛虫发生相关关系的调查[J]. 森林病虫通讯, (2): 34-35. |
孟庆繁. 2006. 人工林在生物多样性保护中的作用[J]. 世界林业研究, 19(5): 1-5. |
彭映辉, 许万景. 1992. 岳麓山的主要植被类型及其分布[J]. 湖南林业科技, 19(4): 59-61. |
彭羽, 刘雪华. 2007. 城市化对植物多样性影响的研究进展[J]. 生物多样性, 15(5): 558-562. |
任海, 彭少麟, 陆宏芳. 2004. 退化生态系统恢复与恢复生态学[J]. 生态学报, 24(8): 1756-1764. |
汤景明, 石冰天, 杜超群. 2007. 不同更新方式对常绿落叶阔叶混交林物种多样性的影响[J]. 湖北林业科技, (5): 1-5. |
田晔林, 陈三茂, 马亮. 2002. 岳麓山生态旅游可持续发展的思路与对策[J]. 北京农学院学报, 17(1): 45-49. |
童丽丽, 汤庚国, 许晓岗. 2007. 城市森林群落结构研究现状及发展评价[J]. 南京林业大学学报:自然科学版, 31(5): 113-116. |
童群益, 徐高福, 王振懂, 等. 2004. 马尾松林阔叶化改造的目标和方法探讨[J]. 浙江林业科技, 24(5): 41-46. |
王贺新, 李根柱, 于冬梅, 等. 2008. 枯枝落叶层对森林天然更新的障碍[J]. 生态学杂志, 27(1): 83-88. |
王巧珍. 2003. 马尾松林下套种阔叶树的物种多样性研究[J]. 福建林业科技, 30(3): 47-50. |
王希华, 宋永昌, 王良衍. 2001. 马尾松林恢复为常绿阔叶林的研究[J]. 生态学杂志, 20(1): 30-32. |
王献溥, 蒋高明. 2002. 广西马尾松林分类, 分布和演替的研究[J]. 植物研究, 22(2): 151-155. |
项文化, 田大伦. 2002. 不同年龄阶段马尾松人工林养分循环的研究[J]. 植物生态学报, 26(1): 89-95. |
杨心兵, 覃逸明, 刘胜祥. 2001. 武汉市马鞍山森林公园马尾松年龄种群结构与分布格局[J]. 华中师范大学学报:自然科学版, 35(2): 209-213. |
张金屯. 2004. 数量生态学[M]. 北京: 科学出版社.
|
张克荣, 刘应迪, 谭淑端, 等. 2009a. 湖南岳麓山植被演替趋势[J]. 生命科学研究, 13(4): 310-313. |
张克荣, 刘应迪, 朱晓文, 等. 2009b. 岳麓山马尾松生态位分析[J]. 生态学杂志, 28(2): 197-202. |
张明. 1997. 岳麓山风景林季相特色植物及其评价[J]. 湖南林业科技, 24(1): 58-62. |
郑景明, 马克平. 2006. 植物群落多样性与可入侵性关系研究进展[J]. 应用生态学报, 17(7): 1338-1343. |
Ashton P M S, Gamage S, Gunatilleke I A U N, et al. 1997. Restoration of a Sri Lankan rainforest:using Caribbean pine Pinus caribaea as a nurse for establishing late-successional tree species[J]. Journal of Applied Ecology, 34(4): 915-925. DOI:10.2307/2405282 |
Balvanera P, Pfisterer A B, Buchmann N, et al. 2006. Quantifying the evidence for biodiversity effects on ecosystem functioning and services[J]. Ecology Letters, 9(10): 1146-1156. DOI:10.1111/j.1461-0248.2006.00963.x |
Brockerhoff E G, Ecroyd C E, Leckie A C, et al. 2003. Diversity and succession of adventive and indigenous vascular understorey plants in Pinus radiata plantation forests in New Zealand[J]. Forest Ecology and Management, 185(3): 307-326. DOI:10.1016/S0378-1127(03)00227-5 |
Brown R L, Peet R K. 2003. Diversity and invasibility of southern Appalachian plant communities[J]. Ecology, 84(1): 32-39. DOI:10.1890/0012-9658(2003)084[0032:DAIOSA]2.0.CO;2 |
Cardinale B J, Srivastava D S, Duffy J E, et al. 2006. Effects of biodiversity on the functioning of trophic groups and ecosystems[J]. Nature, 443: 989-992. DOI:10.1038/nature05202 |
Czerepko J. 2004. Development of vegetation in managed Scots pine(Pinus sylvestris L.) stands in an oak-lime-hornbeam forest habitat[J]. Forest Ecology and Management, 202(1/3): 119-130. |
Lamb D, Erskine P D, Parrotta J A, et al. 2005. Restoration of degraded tropical forest landscapes[J]. Science, 310: 1628-1632. DOI:10.1126/science.1111773 |
Langer E R, Steward G A, Kimberley M O. 2008. Vegetation structure, composition and effect of pine plantation harvesting on riparian buffers in New Zealand[J]. Forest Ecology and Management, 256(5): 949-957. DOI:10.1016/j.foreco.2008.05.052 |
Liu S L, Li X M, Niu L M. 1998. The degradation of soil fertility in pure larch plantations in the northeastern part of China[J]. Ecological Engineering, 10(1): 75-86. DOI:10.1016/S0925-8574(97)10024-6 |
Lugo A E. 1997. The apparent paradox of reestablishing species richness on degraded lands with tree monocultures[J]. Forest Ecology and Management, 99: 9-19. DOI:10.1016/S0378-1127(97)00191-6 |
McPherson E G, Nowak D, Heisler G, et al. 1997. Quantifying urban forest structure, function, and value:the Chicago Urban Forest Climate Project[J]. Urban Ecosystems, 1(1): 49-61. DOI:10.1023/A:1014350822458 |
Oberhauser U. 1997. Secondary forest regeneration beneath pine(Pinus kesiya) plantations in the northern Thai highlands:a chronosequence study[J]. Forest Ecology and Management, 99(1/2): 171-183. |
Scholes M C, Nowicki T E.1998.Effects of pines on soil properties and processes//Richardson D M.Ecology and biogeography of Pinus.Combridge:Cambridge University Press, 341-353.
|