 2010, Vol. 46
2010, Vol. 46文章信息
- 宋于洋, 李明艳, 张文辉
- Song Yuyang, Li Mingyan, Zhang Wenhui
- 生境对古尔班通古特沙漠梭梭种群波动及谱分析推绎的影响
- Effect of Habitat on Population Dynamics and Upscaling Spectrograph of Haloxylon ammodendron Population in Gurbantunggut Desert
- 林业科学, 2010, 46(12): 8-14.
- Scientia Silvae Sinicae, 2010, 46(12): 8-14.
- DOI: 10.11707/j.1001-7488.20101202
-
文章历史
- 收稿日期:2009-08-20
- 修回日期:2010-10-08
-
作者相关文章


2. 石河子大学农学院林学系 石河子 832003
2. Department of Forestry, Agricultural College, Xinjiang Shihezi University Shihezi 832003
种群动态是研究种群数量在时间上和空间上的变动规律,是种群生态学的核心问题。生命表是研究种群数量动态变化和进行种群统计的重要工具,通过编制种群生命表可以揭示种群结构现状,分析种群动态(汪殿蓓等,2007)。植物种群结构和动态是植物个体生存能力与外界环境相互作用的结果(张文辉等,2008)。种群数量变动具有周期性波动,这种植物种群数量的周期性波动存在于所有植被中(伍业钢等,1988; 吴明作等,2000; 刘金福等,2003)。谱分析则是探讨种群天然分布的波动性和年龄更替过程的数学工具,种群天然更新可通过不同龄级的株数分布波动来表现。国内外一些学者运用谱分析揭示了不同植物年龄分布的变动状况,对未来种群的数量波动特征具有重要意义(Stewart,1989;闫淑君等,2002; 张钦弟等,2010)。
荒漠植被群落是一个自然等级系统,在该系统内不同尺度上的主导过程不同。植物的生长不仅与生物学特性有关,也受环境及其他物种相互作用的影响,这种作用具有多尺度特性。当时空尺度发生变化时,生态系统实体和过程的性质也会发生变化,甚至显著变化(辛晓平等,2000),而表现出的断点为标志的尺度域。尺度推绎是生态学理论和应用的核心(张娜,2007),通过尺度推绎可以发现种群波动过程产生非线性变化的临界域。
梭梭(Haloxylon ammodendron)为藜科(Chenopodiaceae)植物,是多年生小乔木或灌木,广泛分布在固定或半固定沙丘。它抗逆性和适应性强,是西北荒漠地区的珍贵植物资源和稳定的沙丘先锋树种(吴进琼等,2007)。以前的学者主要对梭梭的生理生态特性、种群的空间格局和结构等方面(常静等,2006; 宋于洋等,2008)进行了研究。但是古尔班通古特沙漠梭梭种群的数量动态和年龄波动特性研究较少。为此,运用静态生命表和谱分析推绎解释梭梭种群波动过程对环境和尺度依赖性是较好的手段。本文研究目的在于: 1)分析古尔班通古特沙漠梭梭种群在不同环境下的波动特性; 2)揭示在谱分析过程中干旱荒漠地区梭梭种群基波的生态学意义; 3)探讨在特定时间序列下,谱分析推绎过程中梭梭种群波动、种群稳定性的最佳尺度,以确定适宜的取样面积。
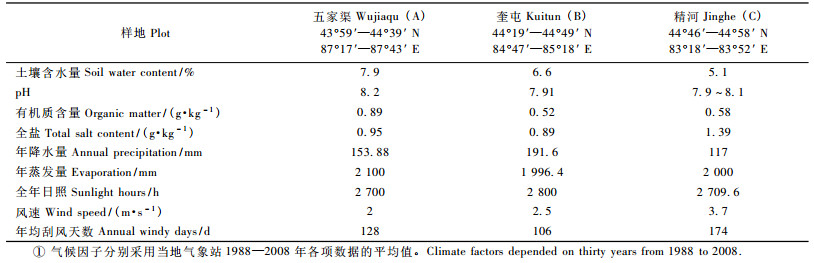
1 研究区概况古尔班通古特沙漠位于44° 15′-46° 50′ N,84°50′-91°20′ E,是中国第二大沙漠,面积4.88万km2。沙漠区为温带干旱荒漠气候,年平均温度5 ~ 5.7 ℃,年降雨量为70 ~ 160 mm。年蒸发量2 000 ~ 2 800 mm,>17.2 m·s-1的大风天数25 ~ 77天。沙漠内部植被覆盖率为15% ~ 30%,在这样的植被条件下,固定和半固定沙丘占整个沙漠面积的96%,沙漠中主要以沙生和耐旱植物为主。
2 研究方法 2.1 调查方法古尔班通古特沙漠景观丰富多样,主要为各种形态的沙垄和一些新月型沙丘链、蜂窝状沙丘,高度10 ~ 30 m。根据古尔班通古特沙漠沙垄外貌的形态特征建立五家渠、奎屯、精河3个典型样地,分别记为样地A,B和C,进行群落调查(表 1)。根据梭梭种群在沙丘间平地和沙垄缓坡最宽处和最长处的分布范围的规律,兼顾3类样地样方大小的一致性,确定每个样地为80 m × 200 m,分成160个10 m × 10 m的小样方,共480个小样方。2008年3-11月,分别对样地植被特征、土壤理化性状和地貌形态进行系统调查和取样并用GPS定位。记录样地中每一株小乔木或灌木的名称、基径、树高、冠幅及生长状况; 每一样地中,在样地的4角及中部分别选取1 m2,分层0 ~ 10 cm和10 ~ 30 cm取土样混匀分析。土壤有机质、养分、全盐含量和pH值,均按常规方法进行测定,土壤含水量采用铝盒取样烘干称量法求出。
|
|
根据相关文献(蔡飞,2000; Knowles et al., 1983; Parker et al., 1984; Rundel,1971),依梭梭过渡径大小分级,径级1 cm,共分为15个径级。根据相关文献(陈远征等,2006),用径级代替年龄编制梭梭种群生命表。
2.3 波谱分析谱分析是Fourier级数的展开。Fourier证明复杂的周期现象可以由不同振幅和相应的谐波组成,写成正弦波形式:
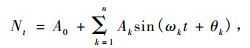
|
式中,A0为周期变化的平均值; Ak为各谐波的振幅,标志其所起的作用大小,其值的差异反映各周期作用大小的差别,k = 1,2,…,n; ωk及θk分别为谐波的频率及相角; Nt为t时刻种群大小(刘彦等,2008)。
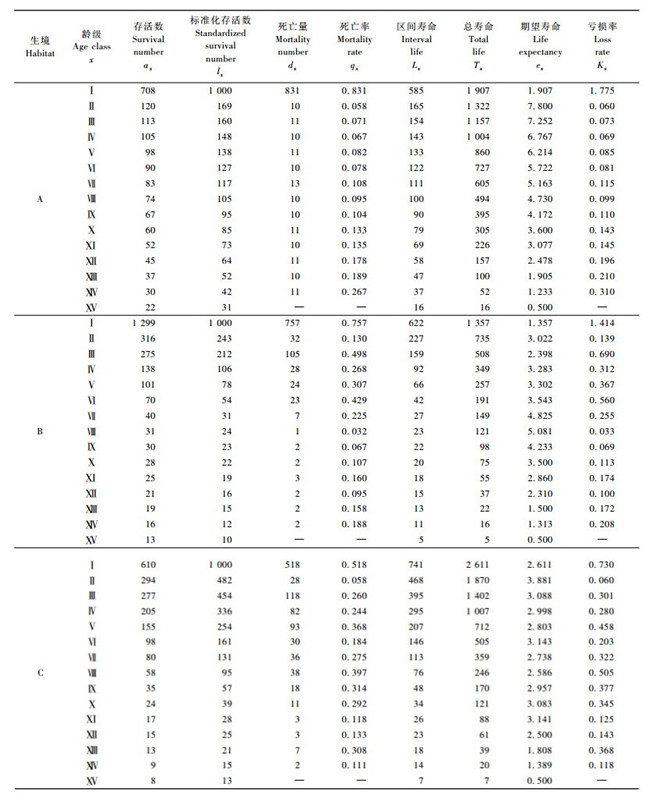
3 结果与分析 3.1 梭梭种群的静态生命表存活曲线是反映种群个体在各龄级的存活状况曲线,根据生命表借助存活个体数量来描述特定龄级死亡率,通过把特定龄组的个体数量相对时间作图而得到的(陈远征等,2006; 江洪,1992)。本研究以径级为横坐标,以标准化存活数(ax)的对数为纵坐标,根据梭梭生命表(表 2),绘制梭梭种群的存活曲线(图 1)。
|
|

|
图 1 不同生境梭梭种群的存活曲线 Figure 1 Survival curves of H. ammodendron populations in different habitats |
由表 2可以看出,梭梭种群静态生命表反映不同生境梭梭种群存亡的基本规律。种群最高死亡率出现的径级、期望寿命的高峰值不同。例如A样地Ⅰ,Ⅶ,Ⅻ,ⅩⅣ径级较高,B样地Ⅰ,Ⅲ,Ⅵ,ⅩⅣ径级较高,C样地Ⅰ,Ⅴ,Ⅷ,ⅩⅣ径级较高。期望寿命反映植物种群的生存能力,不同种群期望寿命的高峰值出现的径级也有差异: A样地是Ⅱ,B样地是Ⅷ,而C样地是Ⅱ径级。梭梭种群在Ⅱ径级以前期望寿命达到最大,表明此阶段种群的生存质量较好,生理活动达到旺盛期; 而后随着径级的增加,期望寿命值陡降,说明在种群的生物学特性和环境条件的双重作用下,梭梭个体的生存力逐渐下降。
3.2 梭梭种群存活曲线及死亡曲线从图 1可以看到,梭梭种群的存活曲线基本接近于Deevey Ⅲ型,而死亡率曲线通常在几个龄级都有较高的死亡率(图 2)。梭梭种子在每年春季萌发,此时沙漠中积雪融化加上春季降水较多,沙地的悬湿沙层给梭梭种子萌发提供了良好的环境条件。5月中旬过后,土壤含水量降低,气温逐渐升高,幼苗死亡率迅速增大,致使Ⅰ,Ⅱ径级受到水分为主的环境因素的强烈筛选而大部分死亡。从图 2可以看出,各样地从Ⅰ到Ⅱ径级死亡率都大幅下降,而随后的各径级则表现不同。A样地为近似直线缓慢上升,B样地和C样地的死亡率曲线呈波浪形,存在4次死亡高峰。不同样地体现不同的生境条件,A样地为土质类型,环境因子基本能够满足梭梭的个体发育,死亡率比较平稳,只是大径级的有所升高,说明后期的生物学特性影响较大; B样地和C样地的环境条件相对较差,死亡率波动明显,说明受环境因子影响较大,特别是水分条件的限制。

|
图 2 不同生境梭梭种群的死亡曲线 Figure 2 Dead curves of H. ammodendron populations in different habitats |
梭梭种群动态的谱分析表明,种群的波动性不是单一周期,而是2个以上周期的迭加,大周期内存在着小周期。各调查林分的Ak值以A1最大,表明梭梭种群数量动态变化受其基波期影响显著。
小周期的谐波A2是基本周期的1/2,周期约为7 cm,对应地径为2 ~ 4 cm,是梭梭由幼苗变成幼树的关键时期; A5是基本周期的1/5,周期约为3 cm,对应地径9 ~ 10 cm,这种周期性正是梭梭种群集中分布的表征; 小周期A7,即地径14 ~ 15 cm与梭梭从成年走向衰老有关。
据调查,土质条件较好的A样地,有机质和土壤含水量相对较高,梭梭生长良好且盖度较高(达43%),各径级数量变化不剧烈。因此,A1值相对其他2个样地偏小,说明A样地受环境因素影响较小,且只存在A4 1个小周期。B样地和C样地属于沙地和砾石土类型,土壤保水性能较差,在长期生存的沙漠环境中,如果遇到几次干旱,则种群数量会明显下降; 反之若遇到几次降水较好的年份,则种群数量将会上升,因而形成小周期波动。
3.4 梭梭种群不同尺度的谱分析推绎把所调查的3个样地数据分别按800,1 200,1 600,2 000,3 200,4 400,5 600,…,15 200 m2划分尺度,共15个尺度进行谱分析推绎(图 4)。不同样地的波动在小尺度变化较大,随着尺度的增大,各样地的波动逐渐趋于稳定,显示出梭梭种群波动的生境异质性特征。

|
图 4 梭梭种群周期性波动的尺度上推绎 Figure 4 Upscaling of Periodic fluctuation of H. ammodendron populations |
A样地在2 000 m2尺度以下表现出A4,A5,即4,3 cm的周期; 2 000 m2尺度以上,只在A4,即4 cm的周期波动,4 400 ~ 5 600 m2尺度以上基本稳定; B样地小周期波期波动与A样地存在差异,800 m2尺度在A4,A5,A7的小周期波动明显,且起作用大小基本相同; 1 200 ~ 3 200 m2尺度谐波A5的作用增强,5 600 ~ 6 800 m2尺度以上基本稳定; C样地在800 ~ 1 200 m2尺度上,A1值较小,从1 600 ~ 3 200 m2尺度,A1值逐渐增大,说明基波的影响逐渐增强,8 000 ~ 9 200 m2尺度以上基本稳定。

|
图 3 不同生境下梭梭种群的周期性波动 Figure 3 Periodic fluctuation of H. ammodendron populations in different habitats |
从不同生境的谱分析尺度推绎来说,随着尺度的增大,小周期波动的作用降低,到达一定尺度时,基波的振幅变化微弱,说明基波对种群波动的作用保持稳定。
4 结论与讨论 4.1 梭梭种群的周期性波动梭梭种群动态过程可以解释为一个周期集中分布的群体代替另一个周期群体向前发展的过程,即梭梭的种群结构和数量变化具有明显的特征。从静态生命表和存活率死亡率曲线分析可知,A样地梭梭种群波动趋势除Ⅰ径级外其余各径级株数分布较均匀,死亡率缓慢上升,表明A样地种群数量稳定; B样地和C样地各径级死亡率不一,有4次死亡高峰,种群波动明显。梭梭种群波动的谱分析还表现出大周期内有小周期的波动,说明其林分的更替是周期循环的,而不是1个连续发展的过程。A样地梭梭种群数量只在A4出现1次小周期波动,而B样地和C样地的梭梭种群出现4次小周期波动,反映出从幼苗转变为成年树的过程中梭梭种群的生长特性和环境对种群数量的调节作用,说明静态生命表及谱分析法在研究梭梭种群波动时,可以反映梭梭种群波动的强烈程度。
植物种群的周期性波动受其生态学特性和环境因素的影响。洪伟等(2004)运用谱分析研究南方红豆杉(Taxus chinensis var.mairei)种群波动时发现存在明显的大周期,其数量动态变化明显受整个生命周期生物学特征的控制。李清河等(2009)对不同坡位荒漠珍稀灌木半日花(Helianthemum songoricum)种群数量动态的谱分析时也发现其种群的数量动态变化受其生命周期中生物学特性的控制趋势极其明显。这些研究说明在环境条件良好的地区,水分、养分条件完全可以满足各径级个体的生长,环境对种群数量动态的调节作用相对小些。在古尔班通古特沙漠,水分等非生物因子制约植物的生长和繁衍(张景光等,2005)。在幼苗期由于气候的波动,常导致绝大部分植株死亡,只有某些年份,因春雨及时滋润才有较多幼苗得以保存。干旱区1个世纪仅有若干次可以成功补充幼苗,使得种群数量呈现波浪式发展。因此,梭梭种群的基波大小不仅受生物学特性的影响,更多地受恶劣环境的影响。这种对基波生物学意义的解释可能与一些学者的不同。比如,在生境条件较好的A样地,各径级数量的波动幅度不如B样地和C样地,说明同一种群的基波在相对较好的环境中主要受生物学特性的控制,而在逆境中更多地受环境因素左右。
本研究未能反映梭梭种群固有周期的长度,主要是所调查的时间系列长度未足够长到表现出基本周期。调查表明,20世纪80年代以前由于人为砍伐、垦荒造成梭梭林面积的锐减,使古尔通古特沙漠南缘30 ~ 50 km范围内已无原始林。此外,由于城市用水和农田面积的扩大致使地下水位严重下降(20世纪70年代浅层水位在1.5 ~ 2 m,80年代末地下水位在5 m左右,现在地下水位在9 ~ 12 m以下),随着地下水位下降根系难以吸收地下水,无法满足较大树体的正常生长。因此,在所调查的样地极少见到过渡径为20 cm左右的大梭梭,致使所调查的时间系列长度不能分辨出大周期的波动长度。
4.2 谱分析的尺度推绎尺度推绎实现了不同时空尺度或组织层次之间的信息转换。在一般情况下,尺度在时间和空间尺度上相似,具有较高的预测性; 如果时间尺度小而空间尺度大,预测性也较高; 但如果时间尺度较大,而空间尺度较小,则预测性较差(Schulze,2000)。由于气候、土壤等自然因素特性的空间变异和干扰机制的普遍存在,使得在该生态系统内,功能要素和空间因素因空间异质性而表现为明显的非线性,相应的系统动态模型(包括尺度推绎模型)往往也要集成空间参变量的非线性功能,因此不能将小尺度上的模型直接用到大尺度上去。通过谱分析尺度推绎发现,梭梭种群的周期性波动特性依赖于尺度大小,并且在不同生境条件下波动稳定时的尺度大小不同。表明在古尔班通古特沙漠相对较好的环境条件下,梭梭种群的周期性波动可在较小的取样面积实现稳定(即样地面积为4 400 ~ 5 600 m2),在相对较差的环境条件下需要较大的取样面积才能实现稳定(即样地面积为4 400 ~ 9 200 m2)。此外,在相同生境条件下小尺度的波动过程对环境变化更为敏感,而大尺度变化较为平稳,如样地C。因此,在进行谱分析时,应针对不同的环境条件确定取样面积的大小,才能分析最佳尺度的周期性波动。
蔡飞. 2000. 杭州西湖山区青冈种群结构和动态的研究[J]. 林业科学, 36(3): 67-72. DOI:10.11707/j.1001-7488.20000312 |
常静, 潘存德, 师瑞锋. 2006. 梭梭-白梭梭群落优势种种群分布格局及其种间关系分析[J]. 新疆农业大学学报, 29(2): 26-29. |
陈远征, 马祥庆, 冯丽贞, 等. 2006. 濒危植物沉水樟的种群生命表和谱分析[J]. 生态学报, 26(12): 4267-4252. DOI:10.3321/j.issn:1000-0933.2006.12.046 |
洪伟, 王新功, 吴承祯, 等. 2004. 濒危植物南方红豆杉种群生命表及谱分析[J]. 应用生态学报, 15(6): 1109-1112. |
江洪. 1992. 云杉种群生态学研究[M]. 北京: 中国林业出版社.
|
李清河, 高婷婷, 李慧卿, 等. 2009. 荒漠珍稀灌木半日花种群数量动态的谱分析[J]. 武汉植物学研究, 27(2): 171-175. |
刘金福, 洪伟. 2003. 格氏拷种群数量动态的谱分析研究[J]. 生物数学学报, 18(3): 357-363. |
刘彦, 余新晓, 岳永杰, 等. 2008. 北京密云水库集水区刺槐种群生活史特征[J]. 北京林业大学学报, 30(增2): 31-35. |
宋萍, 洪伟, 吴承祯, 等. 2008. 珍稀濒危植物桫椤种群生命过程及谱分析[J]. 应用生态学报, 19(12): 2577-2582. |
宋于洋, 刘长青, 赵自玉. 2008. 石河子地区不同生境梭梭种群数量动态分析[J]. 西北植物学报, 28(10): 2118-2124. DOI:10.3321/j.issn:1000-4025.2008.10.029 |
汪殿蓓, 暨淑仪, 陈飞鹏, 等. 2007. 仙湖苏铁种群年龄判断及年龄结构特征[J]. 应用生态学报, 18(3): 476-480. |
吴进琼, 梁机. 2007. 从文献计量分析看我国梭梭的研究现状[J]. 农业图书情报学刊, 19(1): 152-154. |
吴明作, 刘玉萃. 2000. 栓皮栎种群数量动态的谱分析定性[J]. 生态学杂志, 19(4): 23-26. |
伍业钢, 韩进轩. 1988. 阔叶红松林红松种群动态的谱分析[J]. 生态学杂志, 7(1): 19-23. |
辛晓平, 徐斌, 单保庆, 等. 2000. 恢复演替中草地斑块动态及尺度转换分析[J]. 生态学报, 20(7): 587-593. |
许皓, 李彦, 邹婷, 等. 2007. 梭梭生理与个体用水策略对降水改变的响应[J]. 生态学报, 27(12): 5019-5028. DOI:10.3321/j.issn:1000-0933.2007.12.010 |
闫淑君, 洪伟, 吴承祯, 等. 2002. 丝栗栲种群生命过程及谱分析[J]. 应用与环境生物学报, 8(4): 351-355. |
张景光, 王新平, 李新荣, 等. 2005. 荒漠植物生活史对策研究进展与展望[J]. 中国沙漠, 25(3): 306-314. |
张娜. 2007. 生态学中的尺度问题———尺度上推[J]. 生态学报, 27(10): 4252-4263. DOI:10.3321/j.issn:1000-0933.2007.10.037 |
张钦弟, 张金屯, 苏日古嘎, 等. 2010. 庞泉沟自然保护区华北落叶松种群生命表与谱分析[J]. 应用与环境生物学报, 16(1): 1-6. |
张文辉, 卢彦昌, 周建云, 等. 2008. 巴山北坡不同干扰条件下栓皮栎种群结构与动态[J]. 林业科学, 44(7): 11-16. DOI:10.11707/j.1001-7488.20080703 |
Knowles P, Grant M C. 1983. Age and size structure analysis of engelmann spruce, population pine, loadgepole, and limber pine in Cororado[J]. Ecology, 64: 1-9. DOI:10.2307/1937322 |
Parker A J, Peet R K. 1984. Size and age structure of conifer forest[J]. Ecology, 65: 1685-1689. DOI:10.2307/1939148 |
Roland Schulze. 2000. Transcending scales of space and time in impact studies of climate and climate change on agrohydrological responses[J]. Argriculture, Ecosystems and Envirement, 82(1/3): 185-212. |
Rundel P. 1971. Community structure and stability in the giant sequoia groves of the Sierra Nevada, California[J]. American Midland Naturalist, 85(2): 487-192. |
Stewart G H. 1989. The dynamics of old-grow Pseudotsuga forests in the western Cascade Range, Oregon, USA[J]. Vegetation, 82: 79-94. |