 2010, Vol. 46
2010, Vol. 46文章信息
- 王旭, 黄世能, 李家湘, 吴仲民, 周光益, 张守攻, 沈孝清, 刘新科
- Wang Xu, Huang Shineng, Li Jiaxiang, Wu Zhongmin, Zhou Guangyi, Zhang Shougong, Shen Xiaoqing, Liu Xinke
- 冰雪灾害后南岭常绿阔叶林受损优势种萌条特性
- Sprouting Characteristics of Dominant Species in Evergreen Broad-Leaved Forests Damaged by Frozen and Snow Disaster in Nanling Mountain
- 林业科学, 2010, 46(11): 66-72.
- Scientia Silvae Sinicae, 2010, 46(11): 66-72.
-
文章历史
- 收稿日期:2009-11-26
- 修回日期:2010-08-28
-
作者相关文章



2. 中南林业科技大学林学院 长沙 410006;
3. 中国林业科学研究院林业研究所 北京 100091;
4. 广东杨东山十二度水省级自然保护区管理处 乐昌 512200;
5. 广东省龙眼洞林场 广州 510520
2. Central South University of Forestry and Technology Changsha 410006;
3. Research Institute of Forestry, CAF Beijing 100091;
4. Yangdongshan Shierdushui Provincial Nature Reserve Lechang 512200;
5. Longyandong Forest Farm of Guangdong Province Guangzhou 510520
萌条更新是植物适应各种干扰胁迫的有效更新方式(Bond et al., 2001)。在土壤种子库缺乏的情况下,大多数木本植物利用干扰后残留下的枝干活体的萌条迅速恢复森林植被结构和功能,成为木本植物应对干扰的有效适应机制(Bond et al., 1992; 2001; Fagerstrom et al., 1997)。作为一种直接的再生方式,萌条更新在次生林保护、经营和管理中具有重要意义,其生态学意义越来越被研究者所关注(Calvo et al., 2002; Kammesheidt,1998; Pascarella et al., 2000)。国外对干扰后森林萌条更新方面已开展了较多的研究,干扰类型主要包括台风(Walker,1991; Yih et al., 1991; Bellingham et al., 1994)、龙卷风(Glitzenstein et al., 1988; Peterson et al., 1997)、雪灾(Angela et al., 2004)、火灾(Williamson et al., 1986; Kauffman,1991; Sampaio et al., 1993; Miller et al., 1998)、采伐(Gorchov et al., 1993; Pinard et al., 1996)等; 研究内容涉及萌芽在更新中的作用(Simöes et al., 2007)、萌枝的生物学特征、个体生活史策略及影响因素(Bellingham et al., 2000),萌枝在种群、群落和景观水平上的“驻留生态位”效应(Bond et al., 2001)等方面。但我国在这方面的研究较少,仅见个别地点的萌枝初步调查(贺金生等,1998; 何永涛等,2000; 王希华等,2004)以及对一些营林树种萌蘖能力的研究(李景文等,2000; 张纪林等,2004),研究树种主要为杉木(Cunninghamia lanceolata) (马祥庆等,2000; 叶镜中,2007)、杨树(Populus) (方升佐等,2000; 卢景龙等,2001)、水曲柳(Fraxinus mandshurica) (荆涛等,2002)、巨尾桉(Eucalyptus grandis × E.urophylla)(林星华,2001)和刺槐(Robinia pseudoacacia) (董金伟等,2001)等人工造林树种,而对于冰雪灾害受损常绿阔叶林萌条特性的研究鲜见报道。
2008年1—2月,我国南方地区遭受百年罕见的雨雪冰冻天气灾害,南岭常绿阔叶林是此次灾害的重灾区,海拔500 ~ 1 000 m的林区,几乎看不到一株完整的树木(李意德,2008; 肖文发,2008)。灾后森林如何恢复成为人们关注的焦点。受损木萌条的出现成为森林自然恢复的一个重要途径,同时这种大面积受损森林为开展冰雪灾害干扰后森林萌条更新研究提供理想的场所。为此,2008年4月,笔者在广东省乐昌杨东山十二度水省级自然保护区,海拔700 ~ 1 000 m常绿阔叶林内设立样地,开展灾后受损森林萌条特性研究。通过本调查研究,初步揭示南岭受损常绿阔叶林优势种萌条特性,为灾后森林群落的快速恢复提供理论依据,同时也可为亚热带地区生态公益林建设树种筛选提供依据。
1 研究地概况研究地选择广东省杨东山十二度水省级自然保护区内(113° 23′ 09″—113° 29′ 32″ E,25° 22′ 47″—25°11′06″ N),该区气候属于中亚热带季风气候,≥10 ℃年积温为6 386.5 ℃; 无霜期300天左右,年均气温为18.1 ~ 19.9 ℃,1月平均气温7.7 ~9.6 ℃,8月平均气温26.2 ~ 28.1 ℃; 年降水量超过1 700 mm,没有特别明显的干旱季节,雨季在3—8月(王厚麟等,2007)。地貌类型主要是中山地貌,成土母质主要为黑云母花岗岩等,土壤为山地黄壤,表土灰黄色至灰黑色,心土蜡黄色或金黄色。A层土壤厚度为0 ~ 15 cm,土壤密度为0.95 ~ 1.51 g·cm-3,pH在4.5左右,0 ~ 10 cm层土壤有机质量为62 ~106 g·kg-1。林分是经过20世纪80年代初人工采伐后自然更新形成的次生林,森林群落建群种以甜槠(Castanopsis eyrei)、栲(Castanopsis fargesii)、南岭栲(Castanopsis fordii)等锥属植物为主,属亚热带最为典型的常绿阔叶林类型———栲类林(吴征镒,1980)。海拔1 000 m以上为广东松(Pinus kwangtungensis)群落和山顶矮林(祁承经,1992)。
2 研究方法 2.1 调查方法在海拔700 ~ 1 000 m的冰灾受损群落中,按照相邻格子样方的设置方法设4个50 m × 30 m的长方形固定样地,每个样地又分为15个10 m × 10 m的样方。对树木受损情况划分为6类:倒伏、断干、断梢、翻兜、压弯(赵霞等,2008)和死亡,进一步把倒伏、断干、翻蔸类型归为严重受损型,压弯、断梢归为轻度受损型。于2008年11月对样地进行调查,4个样地共调查树木4 047株,其中受损3 648株,产生萌条的2 810株,对样地内胸径(DBH) ≥1 cm木本植物进行每木调查,调查其树高(断干、断梢木以灾后残留高度为树高,压弯、倒伏、翻蔸木以整株长度为树高)、胸径及目前的生长状况,萌条数量及平均高度等。
2.2 分析方法萌条率=样地内产生萌条株数/样地受损活立木数
对样方调查数据采用Excel和SPSS16.0进行统计分析,采用单因子方差分析,Least-significantdifference(LSD) (齐次方差时采用)和Tamhane's T2(非齐次方差时采用)进行平均数方差分析; 相关性分析采用双尾t检验,皮尔逊相关性分析。
3 结果与分析 3.1 群落组成根据调查,4个样地冰灾受损树木隶属于51科87属172个种,群落树种组成以杜鹃花科(Ericaceae)26.61%、壳斗科(Fagaceae) 23.71%、冬青科(Aquifoliaceae) 7.09%、山茶科(Theaceae)7.06%、樟科(Lauraceae) 5.43%、虎皮楠科(Daphniphyllaceae) 4.70%、金缕梅科(Hamamelidaceae)3.96%等植物为表征成分,且这些科内植物在群落构成中优势显著,物种组成以常绿木本植物为主体,是亚热带常绿阔叶林的典型表现。从群落结构组成来看,处于乔木层的主要为甜槠7.49%、雷公青冈(Cyclobalanopsis hui) 7.06%、冬青(Ilex chinensis) 4.80%、南岭栲4.33%、交让木(Daphniphyllum macropodum) 3.20%、广东润楠(Machilus kwangtungensis) 1.90%、小红栲(Castanopsis carlesii) 1.70%、虎皮楠(Daphniphyllum oldhamii)1.50%等,灌木层树种主要为鹿角杜鹃(Rhododendron latoucheae) 10.49%、广东杜鹃(Rhododendron kwangtungense) 9.59%、杜鹃(Rhododendron simsii) 3.20%、格药柃(Eurya muricata) 2.13%、檵木(Loropetalum chinensis)2.06%等。
3.2 不同受损类型对优势种萌条率的影响干扰是刺激树木萌条发生的一个重要因素。从图 1可以看出:倒伏萌条率最高,依次为翻蔸、压弯、断干和断梢,说明在树木根系未受损的情况下,随着损伤程度的增加萌条率提高。轻度受损类型中虎皮楠科树木萌条率最高,杜鹃花科树木次之; 严重受损类型中山茶科树木萌条率最高,壳斗科树木次之。倒伏受损类型中各优势植物科均有萌条发生(在调查的样地中未出现虎皮楠科和金缕梅科),萌条率为100%。其他受损类型中,除翻蔸受损类型外,处于灌木层金缕梅科树木萌条率均为最低; 处于乔木层的树木,虎皮楠科树木萌条率最高,只有在翻蔸受损类型中低于壳斗科树木; 樟科植物萌条率最低,仅在断干受损类型中略高于冬青科树木(高2.27%)。壳斗科和虎皮楠科受损树木萌条率高于冬青科受损树木。这种萌条率的变化可能会改变现有林分组成,如灌木层中,金缕梅科树种可能减少,同时为杜鹃花科和山茶科树木提供更多的生长空间,最终使杜鹃花科与山茶科树种在灌木层的优势更强; 乔木层樟科树木优势度降低,壳斗科和虎皮楠科优势度上升。

|
图 1 不同受损类型对优势种萌条率的影响 Figure 1 Sprouting proportion of damaged dominant species in different damaged modes |
从整体来看,灌木层受损木平均萌条率(79.90%)低于乔木层(87.24%) (图 2),在0.05水平上,灌木层和乔木层层内各优势树种间无显著差异。灌木层鹿角杜鹃平均萌条率最低(72.36%),广东杜鹃平均萌条率最高(93.27%),杜鹃平均萌条数最多(表 1),广东杜鹃平均萌条数最少; 乔木层虎皮楠平均萌条率最高(93.59%),冬青平均萌条率最低(78.57%),小红栲平均萌条数最多,冬青平均萌条数最少。经统计分析:在平均萌条数上,冬青与小红栲(P = 0.003)、广东润楠(P = 0.024)、交让木(P = 0.007)、雷公青冈(P = 0.003)、冬青(P = 0.035),小红栲与甜槠(P =0.005)和南岭栲(P = 0.010)差异显著,其他各乔木层优势种间均无差异; 乔木层在平均萌条高度方面,冬青与广东润楠(P = 0.044)和甜槠(P = 0.010),广东润楠与雷公青冈(P = 0.025),雷公青冈与甜槠(P = 0.006)差异显著。灌木层檵木与格药柃在平均萌条数上有差异(P = 0.032),檵木平均萌条高度与杜鹃(P = 0.01)、广东杜鹃(P = 0.010)和鹿角杜鹃(P = 0.000)有显著差异,其他灌木层优势种间无论在平均萌条数还是平均萌条高度上均无差异。不同冠层间优势树种平均萌条率具有一定的差异,杜鹃与广东润楠(P = 0.04)、虎皮楠(P = 0.025)、交让木(P = 0.042)、小红栲(P = 0.034)间平均萌条率存在显著差异,2层优势种萌条高度间均存在显著差异。说明在短期内灌木层萌条的高生长不如乔木层高生长快。

|
图 2 各受损优势种萌条率 Figure 2 Sprouting proportion of damaged dominant species 1:冬青Ilex chinensis; 2:广东润楠Machilus kwangtungensis; 3:虎皮楠Daphniphyllum oldhamii; 4:交让木Daphnipuyllum macropodium; 5:雷公青冈Cyclobalanopsis hui; 6:南岭栲Castanopsis fordii; 7:甜槠Costanopsis eyrei; 8:小红栲Castanopsis carlesii; 9:杜鹃Rhododendron simsii; 10:格药柃Eurya muricata; 11:广东杜鹃Rhododendron kwangtungense; 12:檵木Loropetalum chinensis; 13:鹿角杜鹃Rhododendron latoucheae. |
|
|
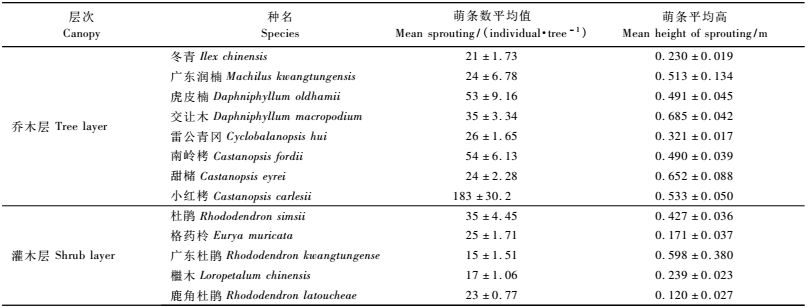
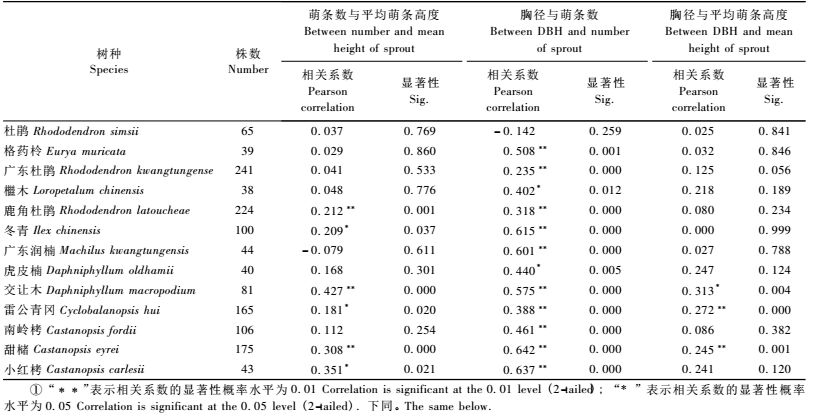
从表 2中可以看出:冰雪灾害受损木中,灌木层优势木鹿角杜鹃外,平均萌条数与平均萌条高度具有显著的相关性,其他种两者间无显著的相关性; 乔木层优势木交让木、甜槠、小红栲和冬青受损后萌条数与平均萌条高度具有显著的相关性,其他种两者间无显著的相关,但广东润楠两者的相关系数为- 0.079,符合萌条数与萌条高生长呈负相关的规律。除杜鹃外,受损木胸径对受损木萌条数影响较大,但受损木胸径对萌条平均高度影响不大,仅有交让木、雷公青冈和甜槠相关性比较明显。从整体来看,胸径对萌条数影响较大,对萌条高度影响不大。
|
|
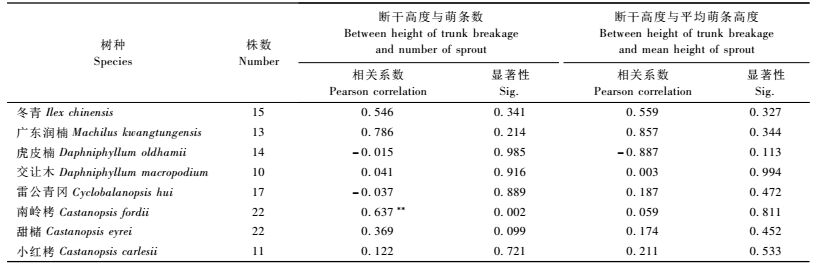
因灌木层以压弯木居多,本文仅对乔木层优势木断干类型进行残留木高度与萌条数和平均萌条高度进行分析。从表 3中可以看出:不同树种间断干高度与萌条数、萌条高度相关性具有较大差异。除南岭栲外,断干高度与萌条数没有显著相关性,虽然广东润楠断干高度与萌条数相关系数较大,但不显著。虎皮楠、雷公青冈断干高度与萌条数和萌条高度呈负相关。在0.05水平上,断干高度与萌条的平均高无显著相关性。乔木层优势木中,虎皮楠和广东润楠断干高度与萌条平均高具有较高的相关性,但表现出相反的相关系数。
|
|
4个样地调查结果表明:调查木90.14%受到不同类型的机械损伤,受损木萌条率高于72.00%,乔木层优势木平均萌条数高于灌木层。在树木根系未受损的情况,机械损伤程度越严重,树木的萌条率越高,同时在笔者调查过程中发现,正常木极少有萌条发生,可见干扰是刺激萌条发生的重要因素。与2000年Angela等(2004)对1998年发生在魁北克特大雪灾害后受损木萌条[其森林主要组成为:糖槭(Acer rubra)、北美红枫( Acer rubrum)、危地马拉松(Pinus spp.)、洋白蜡(Fraxinus pennsylvanica)、水青冈(Fraxinus spp.)、椴木(Tilia americana)、美国榆(Ulmus americana)、加拿大铁木(Ostrya virginiana)等]调查结果25.00%相比,我国南岭地区常绿阔叶林受损木冰雪灾害后萌条率为其2倍以上,也高于Duguay等(2001)在加拿大Montréal调查受损木的萌条率(53.00%)。可见我国南岭地区常绿阔叶林受损木萌条率高于北美落叶林或针阔混交林受损木的萌条率,这也为南岭地区冰雪灾害后受损森林生态系统快速恢复与重建提供保障。
4.2 优势木萌条特性的差异将导致森林结构与组成的变化不同受损优势木萌条率存在差异。倒伏木虽全部有萌条发生,但萌条处于近地表位置,使其失去保持冠层的优势,但对于形成生态系统的复层结构有利。灌木层金缕梅科树木在各受损类型中萌条率基本上最低,乔木层樟科树木萌条率最低; 具体到优势种,灌木层鹿角杜鹃萌条率最低,广东杜鹃萌条率最高,乔木层冬青萌条率最低,虎皮楠萌条率最高,这种现象可能导致干扰后森林结构与组成的变化,进一步影响未来土壤种子库的来源(Pausas et al., 1999; Calvo et al., 2002)。据预测(宋瑞艳等,2008),未来在广东至广西北部和云南中部部分地区,“最大”连续低温日数略有增加; 江西南部、广东东部以及福建西部部分地区降雪量有增加的趋势; 另据在调查过程中与当地林农了解,该区每隔15年左右会有一场雪害发生。如果冰雪灾害在该区发生的频度增加,可能促进萌条率高的树种在群落中的优势更强,从而加速现有群落组成的变化,栲类林的特征更加明显,这也可能是该区形成栲类林的重要因素。
4.3 冰雪灾害后受损木萌条特性(数量和高度)与树木特性(DBH和受损类型)之间相关性分析应做为特例进行研究对萌条数与平均萌条高度分析结果表明:不同树种两者之间相关性不同,鹿角杜鹃、冬青、交让木、雷公青冈、甜槠和小红栲具有相关性,其他优势种相关性不明显。胸径与萌条数具有明显的相关性,这与杨曾奖等(2001)对尾叶桉(Eucalyptus urophylla)伐桩萌芽更新研究结果相一致。除交让木、雷公青冈和甜槠外,胸径与平均萌条高度无明显相关性,这与杨曾奖等(2001)对尾叶桉研究结果不一致。断干高度对萌条的影响,各乔木层优势种表现不同,除南岭栲外,其他优势种萌条数和平均萌条高度无明显相关性。这与叶镜中(2007)对杉木更新的研究结果不同。因此对冰雪灾害后的萌条更新应做为一个特例进行研究。从灾后快速自然恢复方面考虑,萌条率与萌条高度相对萌条数量更为重要。
因此建议在我国亚热带地区森林自然干扰频发区,在人工促进恢复时多选用格药柃、小红栲和交让木等,有利于各层次结构与组成的保持。由于树体的养分制造能力和贮存能力所限,在萌条生长过程中,出现养分竞争,从而导致部分萌条死亡,不能保证养分的集中利用,影响自然快速恢复进程。国内外有关树体对萌条承载力的研究鲜见报道(唐大岳,2004; 张福等,2009),有待进一步加强,这对灾后树体管理和森林经营具有重要意义。虽然1年生萌条中随着时间的推移会有大量萌条死亡(Rijks et al., 1998; Kammesheidt,1999),但大量萌条的出现对短期内提高树冠截流、弥补灾后森林固碳能力等方面发挥着重要生态功能。
董金伟, 杜华兵, 刘呈苓, 等. 2001. 伐桩不同处理对山地刺槐无性更新影响的研究[J]. 山东林业科技, (4): 10-11. |
方升佐, 徐锡增, 吕士行, 等. 2000. 杨树萌芽更新及持续生产力[J]. 南京林业大学学报, 34(4): 43-48. |
何永涛, 曹敏, 唐勇, 等. 2000. 云南省哀牢山中山湿性常绿阔叶林萌生现象的初步研究[J]. 武汉植物学研究, 18(6): 523-527. |
贺金生, 陈伟烈, 刘峰. 1998. 神农架地区米心水青冈萌枝过程的研究[J]. 植物生态学报, 22(5): 385-391. |
荆涛, 马万里, JoniK, 等. 2002. 水曲柳萌芽更新的研究[J]. 北京林业大学学报, 24(4): 12-15. |
李景文, 聂绍荃, 安滨河. 2005. 东北东部林区次生林主要阔叶树种的萌芽更新规律[J]. 林业科学, 41(6): 72-77. DOI:10.11707/j.1001-7488.20050611 |
李意德. 2008. 低温雨雪冰冻灾害后的南岭山脉自然保护区[J]. 林业科学, 44(4): 2-4. DOI:10.11707/j.1001-7488.20080402 |
林武星, 叶功富, 黄金瑞, 等. 1996. 杉木萌芽更新原理及技术述评[J]. 福建林业科技, (2): 19-23. |
林星华. 2001. 巨尾桉二代萌芽更新林分密度调控技术研究[J]. 林业科学研究, 14(3): 283-287. |
卢景龙, 卫金. 2001. 杨树人工林萌芽更新初报[J]. 山西师范大学学报:自然科学版, 15(1): 57-60. |
马祥庆, 刘爱琴. 2001. 杉木免耕萌芽更新生态效果研究[J]. 中南林学院学报, 20(1): 14-18. |
祁承经. 1992. 南岭植被的研究[J]. 中南林学院学报, 12(1): 1-10. |
宋瑞艳, 高学杰, 石英, 等. 2008. 未来我国南方低温雨雪冰冻灾害变化的数值模拟[J]. 气候变化研究进展, 6(4): 352-356. |
唐大岳. 2004. 刺槐萌芽更新及其生长效果研究[J]. 湖北林业科技, 128(2): 23-25. |
王希华, 严晓, 闫恩荣, 等. 天童几种常绿阔叶林优势种在砍伐后萌枝更新的初步研究[J]. 武汉植物学研究, 22(1): 52-57. |
肖文发. 2008. 由亚热带常绿阔叶林雨雪冰冻灾情引发的思考[J]. 林业科学, 44(4): 2-3. DOI:10.11707/j.1001-7488.20080402 |
杨曾奖, 徐大平, 江松远. 2001. 桩径对尾叶桉萌芽更新的影响[J]. 广东林业科技, 17(4): 6-9. |
叶镜中. 2007. 杉木萌芽更新[J]. 南京林业大学学报, 31(2): 1-4. |
张福, 吴昊. 2009. 冀北山地萌生白桦每丛合理保留株数的研究[J]. 安徽农业科学, 37(10): 4666-4667, 4691. DOI:10.3969/j.issn.0517-6611.2009.10.119 |
张纪林, 季永华. 2004. 海岸带杨柳新无性系苗期耐盐性研究[J]. 南京林业大学学报, 28(3): 45-50. |
赵霞, 沈孝清, 黄世能, 等. 2008. 冰雪灾害对杨东山十二度水省自然保护区木本植物机械损伤的初步调查[J]. 林业科学, 44(11): 164-167. DOI:10.3321/j.issn:1001-7488.2008.11.030 |
Angela G, Brommit, Neil Charbonneau, et al. 2004. Crown loss and subsequent branch sprouting of forest trees in response to a major ice storm[J]. Journal of the Torrey Botanical Society, 131(2): 169-176. DOI:10.2307/4126918 |
Bellingham P J, Sparrow A D. 2000. Resprouting as a life history strategy in woody plant communities[J]. Oikos, 89(2): 409-416. DOI:10.1034/j.1600-0706.2000.890224.x |
Bellingham P J, Tanner E V J, Healey J R. 1994. Sprouting of trees in Jamaican montane forests, after a hurricane[J]. Journal of Ecology, 82: 747-758. DOI:10.2307/2261440 |
Bond W J, Cowling R, Michards M. 1992. Competition and coexistence∥ Cowling R M. The ecology of fynbos. Nutrients, fire and diversity. Cape Town: Oxford University Press, 206-225.
|
Bond W J, Midgley J J. 2001. Ecology of sprouting in woody plants: The persistence niche[J]. Trends in Ecology Evolution, 16(1): 45-51. DOI:10.1016/S0169-5347(00)02033-4 |
Calvo L, Tarrega R, Luis E. 2002. Secondary succession after perturbations in a shrubland community[J]. Acta Oecol, 23: 393-404. DOI:10.1016/S1146-609X(02)01164-5 |
Duguay S M, Arh K, Hooper M, et al. 2001. Ice storm damage and early recovery in an old-growth forest[J]. Environment Monitoring and Assessment, 67: 97-108. DOI:10.1023/A:1006464511158 |
Fagerstrom T, Westoby M. 1997. Population dynamics in sessile organisms: some general results from three seemingly different theory lineages[J]. Oikos, 80: 588-594. DOI:10.2307/3546634 |
Glitzenstein J S, Harcombe P A. 1988. Effects of the December 1983 tornado on forest vegetation of the big thicket, southeast Texas, U[J]. S. A. Forest Ecology and Management, 25: 269-290. DOI:10.1016/0378-1127(88)90092-8 |
Gorchov D L, Cornejo F, Ascorra C, et al. 1993. The role of seed dispersal in the natural regeneration of rain forest after strip-cutting in the Peruvian Amazon[J]. Vegetatio, (107 /108): 339-349. |
Kammesheidt L. 1998. The role of tree sprouts in the restoration of stand structure and species diversity in tropical moist forest after slash-andburn agriculture in Eastern Paraguay[J]. Plant Ecology, 139: 155-165. DOI:10.1023/A:1009763402998 |
Kanmeshedt L. 1999. Forest recovery by root suckers and above-ground sprouts after slash-and-burn agriculture, fire and logging in Paraguay and Venezuela[J]. Journal of Tropical Ecology, 15: 143-157. DOI:10.1017/S0266467499000723 |
Kauffman J B. 1991. Survival by sprouting following fire in tropical forests of the eastern Amazon[J]. Biotropica, 23: 219-224. DOI:10.2307/2388198 |
Miller P M, Kauman J B. 1998. Seedling and sprout response to slashand-burn agriculture in a tropical deciduous forest[J]. Biotropica, 30: 538-546. DOI:10.1111/btp.1998.30.issue-4 |
Pascarella J B, Aide T M, Serrano M I, et al. 2000. Land use history regeneration in the Cayey mountains[J]. Puerto Rico Ecosystems, (3): 217-228. |
Pausas J G, Carbó E, Caturla R N, et al. 1999. Post-fore regeneration patterns in the easten Iberian Peninsula[J]. Acta Oecologica, 20(5): 499-508. DOI:10.1016/S1146-609X(00)86617-5 |
Peterson C J, Rebertus A J. 1997. Tornado damage and initial recovery in three adjacent, lowland temperate forests in Missouri[J]. Journal of Vegetation Science, 8: 559-564. DOI:10.2307/3237207 |
Pinard M A, Putz F E. 1996. Retaining forest biomass by reducing logging damage[J]. Biotropica, 28: 278-295. DOI:10.2307/2389193 |
Rijks M H, Malta E J, Zagt R J. 1998. Regeneration through sprout formation in Chlorocardium rodiei (Lauraceae) in Guyana[J]. Journal of Tropical Ecology, 14: 463-475. DOI:10.1017/S0266467498000340 |
Sampaio E V S B, Salcedo I H, Kauman J B. 1993. Effect of different fire severities on coppicing of caatinga vegetation in Serra Talhada, PE, Brazil[J]. Biotropica, 25: 452-460. DOI:10.2307/2388868 |
Simöes C G, Marques M C M. 2007. The role of sprouts in the restoration of Atlantic rainforest in southern Brazil[J]. Restoration Ecology, 15(1): 53-59. DOI:10.1111/rec.2007.15.issue-1 |
Walker L R. 1991. Tree damage and recovery from Hurricane Hugo in Luquillo experimental forest, Puerto Rico[J]. Biotropica, 23: 379-385. DOI:10.2307/2388255 |
Williamson G B, Schatz G E, Avlarado A, et al. 1986. Effects of repeated fires on tropical paramo vegetation[J]. Tropical Ecology, 27: 62-69. |
Yih K, Boucher D H, Vandermeer J H, et al. 1991. Recovery of the rain forest of southeastern Nicaragua after destruction by Hurricane Joan[J]. Biotropica, 23: 106-113. DOI:10.2307/2388295 |