 2010, Vol. 46
2010, Vol. 46文章信息
- 杨洪晓, 王学全, 卢琦, 郭建英, 杨文斌, 褚建民
- Yang Hongxiao, Wang Xuequan, Lu Qi, Guo Jianying, Yang Wenbin, Chu Jianmin
- 行带式柠条锦鸡儿林在内蒙古四子王旗退耕还草工程中的应用
- Optimal Structures of Caragana korshinskii Belts to Make Grassland from Farmland in Siziwang Banner, Inner Mongolia
- 林业科学, 2010, 46(11): 36-42.
- Scientia Silvae Sinicae, 2010, 46(11): 36-42.
-
文章历史
- 收稿日期:2009-04-13
- 修回日期:2010-09-10
-
作者相关文章



2. 中国林业科学研究院荒漠化研究所 北京 100091;
3. 水利部牧区水利科学研究所 呼和浩特 010020;
4. 中国林业科学研究院林业研究所 北京 100091
2. Institute of Destification Studies, Chinese Academy of Forestry Beijing 100091;
3. Institute of Water Resources for Pastoral Areas, Ministry of Water Resources Huhhot 010020;
4. Institute of Forestry, Chinese Academy of Forestry Beijing 100091
退耕还草工程是我国干旱区生态建设的重要举措之一,目的是把一些低产农耕土地修复为草原。然而这并非易事,因为涉及植被的原生演替过程(Clewell et al., 2007)。长期耕作的农田缺乏草原恢复所必需的植物繁殖体,它们必须由邻近草原供给(Graham et al., 1988; 许志信等,2002; 邵新庆等,2005)。即便繁殖体能够由邻近草原传播过来,由于退耕地通常裸露,风力较大,繁殖体不容易停留下来(赵廷宁等,2005; Yang et al., 2006; 赵哈林等,2007)。即便少量繁殖体停留下来并得到萌发机会,由于幼苗处在一种营养贫乏、蒸腾强烈、沙打严重的环境中,很难存活下来(Chan et al., 1988; 于云江等,2003; 韩永伟等,2005; Zhao et al., 2005; 杨树等,2006)。在停止耕作的农田里,用旱生灌木营造防护林可能是突破这些限制的最佳办法(赵哈林等,2007)。大风携带沙尘、枯落物和风播植物的种子或果实,当经过灌木林时,被拦截下来,为草原恢复积聚土壤和繁殖体(刘志民等,2005; 赵廷宁等,2005; 王继和等,2006; Zhao et al., 2007)。而且,由于在灌木林间隙地表风速降低,草本植物还可免受流沙袭击,减少体内水分的无效蒸腾,从而获得更多萌发和生长的机会(刘广菊等,2000; 于云江等,2003; 张华等,2004; Singh et al., 2007)。可是,不少学者对灌木林的作用持有怀疑,认为在干旱草原区,水分相对不足,建造乔木林或灌木林未必合适。如果强行建造,它们必然竞争水分、光照和养分,成为阻碍草原恢复的消极因素,结果树木只能长成“小老头树”,且草原不能正常恢复(曹世雄等,2007)。
不可否认,在干旱区草原是无以代替的地带性植被。这并不意味着那里只能容得下草原,而容不下其他植被(牛建明,2000)。除了草原,还有各种非地带性植被,如局域地段上的湿生植被或森林,等等。即便在地带性地段,当植被成熟度降低时,旱生灌木可侵入并生长多年直到被草本群落自然取代(李新荣等,2001; Rietkerk et al., 2002; 肖洪浪等,2003; 阿拉木萨等,2005)。在退耕农田里营造灌木林并不违背植被的自然分布规律,这些土地的植被相对稀疏,植物的蒸腾强度相对微弱,下渗的雨水可逐年积累并有深层化分布的倾向(肖洪浪等,2003; 王新平等,2005; 田有亮等,2008)。所以,其特定水分条件可暂时允许某些灌木林的生长,为借用灌木促进草原恢复提供可能。当灌木林的密度高于40% ~ 50%时,其防风固沙效果虽好,但耗水量和遮荫面积太大,不利于草原的恢复。可是,若没有灌木的保护,沙化土地迟迟不能固定,草原历经多年不能恢复(Yang et al., 2006; Zhao et al., 2007)。因此,控制造林密度和合理规划灌木的布局是至关重要的。目前,我国学者已解决以低密度行带式灌木林取得良好防风固沙效果的问题(Gustafson et al., 1992; 杨文斌等,2006; 2007; 杨红艳等,2008)。然而,如何把这种结构的灌木林更好地应用于退耕还草工程,对于其中的技术细节,人们尚存不少疑问。
防风固沙与草原恢复是密切相关的过程,然而它们不是完全等同的。为推进干旱区退耕还草的生态建设行动,需要开展更有针对性的研究。因此,以1994年在四子王旗退耕农田营造的行带式灌木林为例,研究12年来灌木林在草原重建方面的作用,以便全面认识其生态作用,并以此为依据,合理规划和构建行带式灌木林,并最大限度发挥其促进草原恢复的作用。
1 研究地区与方法 1.1 研究区概况四子王旗位于内蒙古乌兰察布盟西北部,海拔1 000 ~ 2 100 m。属于典型温带大陆性气候区,气候干燥寒冷,多年平均降水量110 ~ 310 mm,降雨主要集中在7—9月,年潜在蒸发量2 130 ~ 2 750 mm,年均温1 ~ 6 ℃,无霜期90 ~ 120天。地带性植被包括荒漠草原和典型草原,建群种为针茅(Stipa sp.)。此外,羊草(Leymus chinensis)、隐子草(Cleistogenes squarrosa)、狗尾草(Setaria viridis)、蒿类(Artemisia sp.)和多根葱(Allium polyrrhizum)也很常见。四子王旗是以牧业为主、农牧结合的旗县,农作物以春小麦(Triticum aestivum)、莜麦(Avena nuda)、马铃薯(Solanum tuberosum)、油菜(Brassica sp.)、甜菜(Beta rulgaris)为主。风蚀常与农耕为伴,致使农田沙化现象相当严重,土壤肥力连年下降,粮食产量常低于800 kg·hm-2,必须依赖施肥等措施才能维持产量(乌兰察布盟计划委员会,1989)。
自1994年,四子王旗在国内率先展开退耕还草试点研究。为促进草场恢复,在退耕农田尝试用优良旱生灌木柠条锦鸡儿(Caragana korshinskii)建造防护林带。每条林带由2行柠条锦鸡儿构成,行间距约为1 m,当地将这种模式称为“两行一带”。柠条锦鸡儿林带的建造几乎都采用这种模式,但在不同的地块它们的带间距(相邻2条林带之间的距离)可能不同,4 ~ 36 m不等。柠条锦鸡儿长势良好,除生态效益外,经济效益也很明显,嫩枝叶可作为牛羊的饲料,枝条耐刈割,牛羊吃剩的割条可作为薪炭材或工业原料。林带之间没有禁牧,粗大砾石至今可见。尽管如此,林带间隙的植被表现出明显的恢复迹象,冷蒿(Artemisia frigida)、狗尾草、益母草(Leonurus japonicus)、羊草、猪毛菜(Salsola collina)、山苦荬(Ixeris chinensis)、阿尔泰狗哇花(Heteropappus altaicus)、凤毛菊(Saussurea amara)等草原常见种已成功定居。
1.2 野外调查2006—2007年在四子王旗退耕还草实验基地开展调查,在当地林业局的协助下找到1994年以带间距12,20和36 m建造的柠条锦鸡儿防护林带,林带为南北走向,柠条锦鸡儿约2 m高。用1 m × 1 m的样方对林带间隙的草本群落进行调查,在每个样方内均匀布设100个样点,样点面积可以忽略。根据这100个样点上是否有植物出现以及相应植物种类,计算每个样方的植物盖度和物种数目,植物盖度等于含有植物的样点总数除以100。对于带间距为12 m的情形,从距东侧林带1 m处开始调查,每隔2 m作为1个梯度,直到距东侧林带11 m且距西侧林带1 m处,共6个梯度。对于带间距为20 m的情形,从距东侧林带1 m处开始调查,每隔2 m作为1个梯度,直到距东侧林带19 m且距西侧林带1 m处,共10个梯度。对于带间距为36 m的情形,从距东侧林带1 m处开始,每隔7 m作为1个梯度,直到距东侧林带35 m且距西侧林带1 m处,共6个梯度。在每个梯度设置3个重复样方,共采集66个样方。
1.3 数据分析1) 对以样方为单元的植物盖度和物种数目进行双因素方差分析,以初步判断带间距和到林带的距离是否影响样方的植物盖度和物种数目。根据统计学原理,确定以带间距为第1因素,以到林带的距离为第2因素,第2因素是基于第1因素的系统分组,而非交叉分组。而且,由于与第2因素相关联的土壤条件是不确定因素,选用系统分组型的随机效应模型进行双因素方差分析(刘来福等,1988; 骆建霞等,2001)。以EXCEL为辅助工具,以统计学原理为指导,进行离差平方和的逐级分解和F检验。
2) 分析在每种带间距条件下出现于林带间隙的草本植物及其生态位宽度。这里采用了Levins生态位宽度(W),用来反映一个物种在特定带间距条件下的广泛或普遍程度(张金屯,2004; 王祥福等,2008)。一般而言,生态位宽度越大,表明适合该物种生活的空间越宽广,在这种条件下该物种越普遍或越广泛。
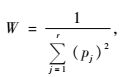
|
(1) |
式中,W为Levins生态位宽度,pj是特定物种在第j个资源状态或样方中的盖度同该物种在所有样方中盖度总和的比值,r表示资源状态或样方的数目。
3) 分别以线性模型和单峰模型拟合样方到林的距离同样方的植物盖度或物种数目的关系。因为林带间隙所受影响来自两侧林带,对于任一样方都有2个距离参数,不适合建立一元模型,所以用公式2对每个样方的距离进行转化,转化后的距离称为折合距离。该公式的假定条件是: a)林带对东西两侧的影响是无差别的; b)林带对林带间隙单一环境因子的影响力同各样方到林带的距离成反比。根据公式,距某侧林带为x,距另一侧林带为y的样方,相当于距一侧林带为z而另一侧没有林带的情况。随后,以折合距离z为自变量,以植物盖度和物种数目为因变量分别进行直线拟合和单峰曲线拟合。单峰曲线是一类曲线,包含多种具体形式,这里尝试以最常用的高斯模型进行拟合。拟合过程在数学软件Origin Pro 8.0中完成。
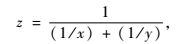
|
(2) |
式中,x和y分别为样方到东西两侧林带的距离,z为折合距离,相当于当一侧有林带而另一侧没林带时样方到林带的距离。
2 结果与分析 2.1 描述性统计关于植物盖度的双因素方差分析表明,林带间距和在林带间隙中所处的位置都显著影响植物盖度(P<0.01)。当林带间距为20 m时,植物盖度是最高的,平均盖度达到32%,明显高于带间距为12 m和36 m的情形(图 1)。关于物种数目的双因素方差分析表明,林带间距能显著影响物种数目(P<0.01),但在林带间隙中所处的位置对物种数目的影响未达到显著程度(P>0.05)。当带间距为12 m和20 m时,植物的物种数目是比较多的,平均多于8个; 当林带间距为36 m时,植被的物种数目是比较少的,平均值约为7个(图 1)。因为植物盖度和物种多样性可作为衡量植被恢复程度的重要指标(高尚武等,1998; 杨洪晓等,2006; Yang et al., 2006),所以初步判断当带间距过大或过小时,对林带间隙的植被恢复都是不利的。

|
图 1 宽度不同的林带间隙的植物盖度和物种数目 Figure 1 Vegetation cover and species number of different intervals in different sizes |
草原地区的一些常见物种,如羊草、蒿类、阿尔泰狗哇花、瓣蕊唐松草(Thalictrum petaloideum)、萹蓄豆(Pocockia ruthenia)、草木樨(Melilotus suaveolens)、田旋花(Convolvulus arvensis)等20多种草本植物已完成定居过程,但当地草原的建群种针茅还没定居下来,可能是土壤尚不适合针茅的缘故(表 1)。在3种带间距条件下,林带间隙草本群落的物种组成和生态位宽度有一定差异(表 1)。总体而言,多数物种在带间距为20 m时表现出较大的生态位宽度,只有少数物种在带间距为12 m或36 m时表现出较大的生态位宽度。这表明,当带间距为20 m时林带间隙的环境适合较多植物的定居与繁衍,然而当带间距为12 m或36 m时林带间隙的环境只适合少数物种的定居与发展。
|
|

比较样方到林带的距离同样方盖度、物种数目的拟合关系,可以发现用高斯模型得到的拟合结果明显优于用线性模型得到拟合结果,其中植物盖度高斯拟合曲线的最大值出现在折合距离为4 m处(图 2),物种数目高斯拟合曲线的最大值出现在折合距离为3.57 m处(图 3),两者比较接近。这些拟合曲线都达到极显著水平(P = 0.001),而线性模型未达到显著水平(P>0.05) (图 2,3),这表明柠条锦鸡儿林带确实影响林带间隙的植被恢复过程,这种影响同各样方到林带的距离有关。根据这些拟合曲线,可以判断: 1)当与柠条锦鸡儿林带的折合距离大约为4 m时,林带对植被恢复的促进作用是最明显的; 2)当与柠条锦鸡儿林的折合距离不足4 m时,植被恢复受到的促进作用反而减弱; 3)当与柠条锦鸡儿林的折合距离超出4 m时,植被恢复受到的促进作用也会减弱; 4)当与柠条锦鸡儿林的折合距离大于7 m时,林带对植被恢复的促进作用将消失。

|
图 2 植物盖度同样方到林带的折合距离的关系 Figure 2 Regression relationships of vegetation cover with modified distances from a shrub belt 线性模型(Linear model) : v = 22.33-0.085z (F = 0.02<F0.05 (1,64) = 3.99,p = 0.89)。 高斯模型(Gaussian model) : 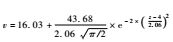 (F = 90.57>F0.001 (4,62) = 5.28,p<0.001)。上述两式中,v表示植物盖度,z表示到单一林带的折合距离。In the formulae,thevariable v represents values of vegetation coverage,and z represents modified distances to a single belt (F = 90.57>F0.001 (4,62) = 5.28,p<0.001)。上述两式中,v表示植物盖度,z表示到单一林带的折合距离。In the formulae,thevariable v represents values of vegetation coverage,and z represents modified distances to a single belt |

|
图 3 物种数目同样方到林带的折合距离的关系 Figure 3 Regression relationships of species number with modified distances from a shrub belt 线性模型(Linear model) : s = 8.19-0.041z (F = 0.16<F0.05 (1,64) = 3.99,p = 0.69)。 高斯模型(Gaussian model) : 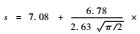 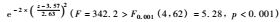 。上述2式中,s表示物种数目,z表示到单一林带的折合距离。In theformulae,the variable s represents species number,and z representsmodified distances to a single belt. 。上述2式中,s表示物种数目,z表示到单一林带的折合距离。In theformulae,the variable s represents species number,and z representsmodified distances to a single belt. |
既然农田的植被恢复效果同它们到林带的距离有关,便可通过优化灌木林的结构,发挥林带的积极影响并减轻消极影响。根据高斯拟合曲线,比较明智的做法是让林带间隙内最难防护地点的折合距离z大于4 m且小于7 m。假定林带间距为d,依据折合距离的计算公式2可以算出,当x和y同时取d/2时,折合距离z取得最大值d/4,即这里最难防护。只有让d在16 ~ 28 m之间取值,折合距离z的最大值才会在4 ~ 7 m之间变动。所以,在建设行带式柠条锦鸡儿防护林时,最好让林带间距d介于16 ~ 28m。如果林带间距小于16 m,那么最难防护地点的折合距离小于4 m,会浪费林带的正面促进作用,无效增加单位面积上负面效应。当林带间距为12 m时,林带间隙的植被恢复效果反倒不如林带间距为20 m时好。如果林带间距大于28 m,那么最难防护地点的折合距离大于7 m,这样就会有部分林带间隙得不到防护。当林带间距为36 m时,林带间隙的植被恢复效果也不如林带间距为20 m时好。根据这些预测,如果采用16 ~ 28 m的带间距,几十年后草原能较好地恢复起来。那时柠条锦鸡儿可能因水分不足而死亡,但它们已完成促进草原恢复的历史使命。也许在柠条锦鸡儿死后会留下部分空地,但周围草本容易扩散过去并修复它们,不会让它们再度出现严重的退化或沙化。
3 讨论 3.1 林带间隙草原恢复的空间差异在研究中,采用草本植物的多样性和盖度作为评价草原恢复进程的指标。虽然没有采用光照、水分、养分、土壤种子库、风沙流等因素,却未把这些因素排除于考虑范围,这种方法不违背国内外学者对此类研究的通常做法(许志信等,2002; 李裕元等,2004; Yang et al., 2006; Zhao et al., 2007; Zhang et al., 2007)。基于林带间隙草本群落在多样性和盖度方面的空间差异,可以认识草本群落恢复进程的空间差异。造成这种差异的决定性因素只能是两侧的林带。当林带出现于退耕地后,可改变周围的条件,如光照、风、土壤水分、土壤养分、风沙流等(向开馥,1991; 刘广菊等,2000; 张华等,2004; 宗萍萍等,2005; 王继和等,2006)。根据文献,可以知道这些条件的变化与退耕地到林带的距离有密切关系,在距林带远近程度不同的地方这些环境条件发生变化的性质和幅度是不同的(向开馥,1991; 杨文斌等,2006; 杨红艳等,2008)。由于这些条件的空间差异,植物繁殖体的输入、萌发、矿质代谢、水分代谢、光合作用、呼吸作用、生长和存活等生态或生理过程也表现出空间差异,表现在群落层次上就是物种多样性和植物盖度的空间差异,即在林带间隙的不同部位,物种多样性和盖度是不同的(于云江等,2003; 姜丽娜等,2009)。在靠林带很近的地方,风力小,有利于繁殖体和矿物质的积累和数量增加,但是土壤水分容易被柠条锦鸡儿夺去,阳光容易受到柠条锦鸡儿的遮蔽,不利于草本植物的生长。在靠林带很远的地方,尽管阳光相对充足,土壤水分相对丰富,但是风力大,繁殖体和矿物质不易停留和积聚,蒸腾作用强烈,制约草本植物的定居。只有在距离林带为最适数值时,繁殖体、土壤水分和阳光均不少,即各种条件都相对适宜。依据Liebig最小因子定律,这时制约草本群落恢复的最小因子优于其他任何距离值时,所以草本群落的恢复程度最好,草本植物的多样性和盖度最高。
3.2 线性模型对单峰模型值得关注的是为什么行带式柠条锦鸡儿林对植被恢复的影响符合单峰模型而不是线性模型。柠条锦鸡儿林对植被恢复的影响包括2个方面,即正面的促进效应和负面的抑制效应。柠条锦鸡儿林截留从空中或地面输入的种实等繁殖体,还可通过降低地表风速把繁殖体、枯落物和细土保持在林带周围,为土壤发育和植物的定居与繁衍创造有利条件(赵廷宁等,2005; 赵哈林等,2007; Zhao et al., 2007)。另外,林带可以降低风速,使植物的蒸腾作用减弱,有利于提高周围植物的水分利用效率和生长速率(向开馥,1991; 刘广菊等,2000; 于云江等,2003; 杨树等,2006)。这些集中体现为对草原恢复的促进作用。然而,因为柠条锦鸡儿林可以争夺周围的土壤水分,可以对周围造成遮荫,所以还会对林带间隙的植被恢复产生抑制作用(王志强等,2005; 王新平等,2005)。这2个方面的作用是相反的,然而都与到林带的距离有关,距林带越近这些作用越强,距林带越远这些作用越弱(向开馥,1991)。它们随距离增加而减弱的步调未必一致,结果导致它们的综合作用遵循单峰变化。当距林带很近时,这些正面促进作用很强,可是这些负面抑制作用也很强,结果它们的综合作用未必是明显的促进作用; 当距林带很远时,这些负面抑制作用很弱,可是这些正面促进作用也很弱,结果它们的综合作用也不可能是明显的促进作用; 当与林带保持某个适当的距离时,尽管促进作用降低了,但是抑制作用更低,结果2方面综合起来的作用却是相当明显的促进作用。这里只尝试了高斯模型,或许用其他形式的单峰模型也能得到类似结果,甚至更好的结果。然而,不管用什么具体形式,单峰模型可能是最好的模型,这是认识促草型灌木林的关键。
3.3 合理性与局限性这项研究的关键是运用公式2,将林带间隙内各样方到两侧林带的距离转化为折合距离z。该公式的假定条件是: 1)林带对东西两侧的影响是无差别的,2)林带对林带间隙单一环境因子的影响力与各地点到林带的距离成反比,也就是说是以距离的倒数作为权重的。这2个条件都是根据经验得出的假定,未必恰好满足(向开馥,1991)。首先,林带对两侧的影响可能相似,但未必完全相同。其次,林带对两侧的影响主要表现为水分竞争、养分竞争、枝叶遮荫等负面影响,还有降低风速、减少地表风蚀等正面影响(陈炳浩等,2003; 宗萍萍等,2005; 赵哈林等,2007)。它们都随距离的增加而急速下降,用距离的倒数作为权重可能近似地符合这种趋势,但不能确定真实规律是否如此(杨文斌等,2006; 2007; 杨红艳,2008)。在无法精确把握客观规律的时候,尽量接近客观规律也是科学研究的追求目标。另外,林带对环境因子的影响同它们对林带间隙植被恢复的影响是不同的,林带对各项环境因子的影响是直接的,可以用距离的倒数作为权重近似反映它们的影响力,但对植被的影响主要是间接的、综合的和可累积的。林带通过影响水分、光照、风力等环境因子而影响植被,而且这些因子要综合起来才能对植被产生作用,这种综合不仅是在同一时刻上的综合,而且是在过去12年间不断地累积和综合。当前林带间隙的植被是过去12年里不断发展的结果,所以林带对林带间隙草本群落的影响更复杂,不同于对各项环境因子的影响方式。
4 结论行带式柠条锦鸡儿林对草原恢复过程的影响是复杂的,既有积极作用,也有消极作用。当退耕地与林带的距离保持适当时,其恢复过程可受到良好促进。相反,要么积极效应极其微弱,要么消极效应严重抑制积极效应,不利于发挥它们对草原的修复作用。
这一认识对改进四子王旗等草原地带的退耕还草措施具有参考价值。以促草型柠条锦鸡儿林来说,在营造林带时应把带间距控制在16 ~ 28 m,否则不能有效发挥它们对于草原恢复过程的促进作用。除了柠条锦鸡儿林带,还有其他灌木林带,可能遵循类似规律。
阿拉木萨, 蒋德明, 骆永明. 2005. 半干旱区人工固沙灌丛发育过程土壤水分及水量平衡研究[J]. 水土保持学报, 19(4): 107-110. |
曹世雄, 陈军, 陈莉, 等. 2007. 退耕还林项目对陕北地区自然与社会的影响[J]. 中国农业科学, 40(5): 972-979. |
陈炳浩, 郝玉光, 陈永富. 2003. 乌兰布和沙区区域性防护林体系气候生态效益评价的研究[J]. 林业科学研究, 16(1): 63-68. |
高尚武, 王葆芳, 朱灵益, 等. 1998. 中国沙质荒漠化土地监测评价指标体系[J]. 林业科学, 34(2): 1-10. |
韩永伟, 韩建国, 张蕴薇, 等. 2005. 农牧交错带退耕还草地土壤风蚀影响因子分析[J]. 生态环境, 14(3): 382-386. |
姜丽娜, 杨文斌, 卢琦, 等. 2009. 低覆盖度柠条固沙林不同配置对植被修复的影响[J]. 干旱区资源与环境, 23(2): 180-185. |
李新荣, 马凤云, 龙立群, 等. 2001. 沙坡头地区固沙植被土壤水分动态研究[J]. 中国沙漠, 21(3): 217-222. |
李裕元, 邵明安. 2004. 子午岭植被自然恢复过程中植物多样性的变化[J]. 生态学报, 24(2): 252-260. DOI:10.17521/cjpe.2004.0037 |
刘广菊, 胡光, 曲海红. 2000. 半干旱风沙区疏林式草牧场防护林气象效应[J]. 东北林业大学学报, 28(5): 83-86. |
刘来福, 程书肖. 1988. 生物统计学[M]. 北京: 北京师范大学出版社: 259-272.
|
刘志民, 蒋德明, 阎巧玲, 等. 2005. 科尔沁草原主要草地植物传播生物学简析[J]. 草业学报, 14(6): 23-33. |
骆建霞, 孙建设. 2001. 园艺植物科学研究导论[M]. 北京: 中国农业出版社: 204-212.
|
牛建明. 2000. 内蒙古主要植被类型与气候因子关系的研究[J]. 应用生态学报, 11(1): 47-52. |
邵新庆, 王堃, 吕进英. 2005. 华北农牧交错带退化草地土壤种子库动态变化[J]. 草业科学, 22(11): 8-12. DOI:10.3969/j.issn.1001-0629.2005.11.002 |
田有亮, 何炎红, 郭连生. 2008. 乌兰布和沙漠东北部土壤水分植被承载力[J]. 林业科学, 44(9): 13-19. DOI:10.11707/j.1001-7488.20080903 |
王继和, 马全林, 刘虎俊, 等. 2006. 干旱区沙漠化土地逆转植被的防风固沙效益研究[J]. 中国沙漠, 26(6): 903-909. |
王祥福, 郭泉水, 巴哈尔古丽, 等. 2008. 崖柏群落优势乔木种群生态位[J]. 林业科学, 44(4): 6-13. DOI:10.11707/j.1001-7488.20080404 |
王新平, 李新荣, 肖洪浪, 等. 2005. 干旱半干旱地区人工固沙灌木林生态系统演变特征[J]. 生态学报, 25(8): 1974-1980. |
王志强, 刘宝元, 王晓兰. 2005. 黄土高原半干旱区天然锦鸡儿灌丛对土壤水分的影响[J]. 地理研究, 24(1): 113-120. |
乌兰察布盟计划委员会. 1989. 乌兰察布盟国土资源[M]. 呼和浩特: 内蒙古人民出版社.
|
伍光和, 王文瑞. 2002. 地域分异规律与北方农牧交错带的退耕还林还草[J]. 中国沙漠, 22(5): 439-442. |
向开馥. 1991. 防护林学[M]. 哈尔滨: 东北林业大学出版社.
|
肖洪浪, 李新荣, 段争虎, 等. 2003. 流沙固定过程中土壤-植被系统演变[J]. 中国沙漠, 23(6): 605-611. |
许志信, 李永强, 额尔德尼, 等. 2002. 草原弃耕地植物群落特征和植被演替情况的调查研究[J]. 内蒙古草业, 14(3): 10-13. |
杨红艳, 戴晟懋, 乐林, 等. 2008. 不同分布格局低覆盖度油蒿群丛防风效果[J]. 林业科学, 44(5): 11-16. DOI:10.11707/j.1001-7488.20080505 |
杨洪晓, 卢琦, 吴波, 等. 2006. 青海共和盆地沙化土地生态修复效果的研究[J]. 中国水土保持科学, 4(2): 7-12. |
杨树, 温雨金, 刘鸿雁. 2006. 内蒙古中部地区退耕还林还草后植被与土壤性状的变化[J]. 水土保持研究, 13(4): 143-147. |
杨文斌, 丁国栋, 王晶莹, 等. 2006. 行带式柠条固沙林防风效果[J]. 生态学报, 26(12): 4106-4113. DOI:10.3321/j.issn:1000-0933.2006.12.025 |
杨文斌, 卢琦, 吴波, 等. 2007. 杨树固沙林密度、配置与林木生长过程的关系[J]. 林业科学, 43(8): 54-59. |
于云江, 史培军, 鲁春霞, 等. 2003. 不同风沙条件对几种植物生态生理特征的影响[J]. 植物生态学报, 27(1): 53-58. DOI:10.17521/cjpe.2003.0008 |
张华, 李锋瑞, 伏乾科, 等. 2004. 沙质草地植被防风抗蚀生态效应的野外观测研究[J]. 环境科学, 25(2): 119-124. |
张金屯. 2004. 数量生态学[M]. 北京: 科学出版社: 113.
|
赵哈林, 苏永中, 张华, 等. 2007. 灌丛对流动沙地土壤特性和草本植物的影响[J]. 中国沙漠, 27(3): 385-390. |
赵廷宁, 曹子龙, 郑翠玲, 等. 2005. 平行高立式沙障对严重沙化草地植被及土壤种子库的影响[J]. 北京林业大学学报, 27(2): 34-37. |
宗萍萍, 鲍玉海, 杨吉华, 等. 2005. 黄泛沙地小网格农田防护林网防护效应的研究[J]. 水土保持学报, 19(6): 110-113. |
Chan K Y, Bellotti W D, Roberts W P. 1988. Changes in surface soil properties of vertisols under dryland cropping in a semiarid environment[J]. Australian Journal of Soil Research, 26(3): 509-518. DOI:10.1071/SR9880509 |
Clewell A F, Aronson J. 2007. Ecological restoration: Principles, values, and structure of an emerging profession[M]. Washington D C: Island Press: 19-32.
|
Graham D J, Hutchings M J. 1988. Estimation of the seed bank of a chalk grassland ley established on former arable land[J]. Journal of Applied Ecology, 25: 241-252. DOI:10.2307/2403622 |
Gustafson E J, Parker G R. 1992. Relationships between landcover proportion and indices of landscape spatial pattern[J]. Landscape Ecology, 7(2): 101-110. DOI:10.1007/BF02418941 |
Rietkerk M, Boerlijst M C, van Langevelde F, et al. 2002. Selforganization of vegetation in arid ecosystems[J]. American Naturalist, 160(4): 524-530. |
Singh G, Mutha S, Bala N. 2007. Effect of tree density on productivity of a Prosopis cineraria agroforestry system in North Western India[J]. Journal of Arid Environments, 70(1): 152-163. DOI:10.1016/j.jaridenv.2006.12.003 |
Yang H, Lu Q, Wu B, et al. 2006. Vegetation diversity and its application in sandy desert revegetation on Tibetan Plateau[J]. Journal of Arid Environments, 65(4): 619-631. DOI:10.1016/j.jaridenv.2005.08.010 |
Zhang J T, Chen T G. 2007. Effects of mixed Hippophae rhamnoides on community and soil in planted forests in the Eastern Loess Plateau, China[J]. Ecological Engineering, 31(2): 115-121. DOI:10.1016/j.ecoleng.2007.06.003 |
Zhao H L, Zhou R L, Su Y Z, et al. 2007. Shrub facilitation of desert land restoration in the Horqin Sand Land of Inner Mongolia[J]. Ecological Engineering, 31(1): 1-8. DOI:10.1016/j.ecoleng.2007.04.010 |
Zhao W Z, Xiao H L, Liu Z M, et al. 2005. Soil degradation and restoration as affected by land use change in the semiarid Bashang area, northern China[J]. Catena, 59(2): 173-186. DOI:10.1016/j.catena.2004.06.004 |