 2010, Vol. 46
2010, Vol. 46文章信息
- 刘华, 臧润国, 丁易, 张炜银, 郭仲军, 白志强, 刘双成
- Liu Hua, Zang Runguo, Ding Yi, Zhang Weiyin, Guo Zhongjun, Bai Zhiqiang, Liu Shuangcheng
- 天山西部新疆野苹果种群特征
- Population Characteristics of Malus sieversii in the West Part of Tianshan Mountains, Xinjiang
- 林业科学, 2010, 46(11): 1-7.
- Scientia Silvae Sinicae, 2010, 46(11): 1-7.
-
文章历史
- 收稿日期:2009-12-11
- 修回日期:2010-02-08
-
作者相关文章



2. 中国林业科学研究院森林生态环境与保护研究所国家林业局森林生态与环境重点实验室 北京 100091;
3. 新疆林业科学研究院森林生态研究所 乌鲁木齐 830000;
4. 天西林业局巩留林场 巩留 835401
2. Key Laboratory of Forest Ecology and Environment, the State Forestry Administration Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry Beijing 100091;
3. Institute of Forest Ecology, Xinjiang Academy of Forestry Science Urumuqi 830000;
4. Forest Farm of Gongliu County, Forestry Bureau of Tianxi Gongliu 835401
新疆野苹果(Malus sieversii)又称塞威士苹果,系蔷薇科(Rosaceae)苹果属(Malus)落叶小乔木,为第三纪孑遗植物之一(傅立国,1992)。它不仅具有较高的经济价值,而且对于研究第三纪植物区系起源以及中国植物区系等具有很高的科研价值。该种喜温暖湿润环境,耐旱力较差,目前在我国仅分布于新疆天山西部的伊犁河谷地区。新疆野苹果群落是新疆天山野果林最主要的群落类型,其分布面积广(占野果林总面积的90%)、类型独特、并蕴含多种野生果树资源,是世界野果基因库的重要组成部分(Christopher et al., 2009),常成片分布在海拔1 000 ~ 1 700 m的中、低山地带具有深厚黄土层的缓斜阴坡和沟谷的逆温层内(佘定域,1995; 刘立诚等,1999; 林培钧等,2002; 羊海军等,2003)。自20世纪50年代以来,新疆野苹果林受到高强度开发利用的影响,如在林中从事放牧、农垦生产、大规模野苹果采收加工、旅游等活动,导致新疆野苹果林从1959年的9 330 hm2下降到目前的不足5 000 hm2,且分布范围急剧缩小,分布区水土流失与山体滑坡现象加剧。更为严重的是,自1999年以来新疆野苹果林受苹果小吉丁虫(Agrilus mali)蔓延的危害,大量的新疆野苹果树木死亡。目前新疆野苹果已被列为我国具有国际意义的生物多样性优先保护物种和中国濒危二级重点保护植物(傅立国,1992)。
种群结构和分布格局都是植物本身生物学特性与环境因素相互作用的结果(Crawly,1986),二者不仅可以反映种群的现实状况和演变的历史与发展趋势(Münzbergová,2004; Nagashima et al., 2009),也是理解植物群落结构组成与动态变化机制的关键(Condit et al., 2000; Hou et al., 2004)。新疆野苹果具有重要的科学研究和种质资源保存意义以及巨大的潜在利用价值,对当地生态环境和社会发展起着重要的作用,越来越引起社会各界的广泛关注。目前,对天山西部新疆野苹果林的研究多集中在描述植物资源分布、形态变异、细胞学(Zhou et al., 2000; Luby et al., 2002; Volk et al., 2005; Zhang et al., 2007; Yan et al., 2008; Christopher et al., 2009; 姜彦成等,2002; 羊海军等,2003; 侯博等,2005; 崔大方等,2006; 冯涛等,2006; 王磊等,2006; 苏馨花等,2007)和病虫害致危原因(阎国荣等,2001; 刘爱华等,2007)等方面,有关其种群结构和资源现状的环境解释还少见详细报道。本研究利用野外调查数据,从种群结构及其分布环境角度分析野苹果种群特征,以期了解新疆野苹果种群基本结构特征与资源现状,为新疆野苹果种群的有效保护和合理开发利用提供依据。
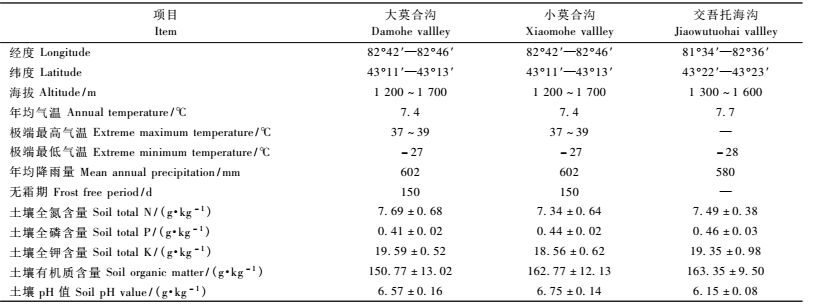
1 研究区概况调查样地位于新疆野苹果天然林3个典型分布区新疆伊犁州的大莫合沟、小莫合沟和交吾托海沟。3个新疆野苹果种群分布区均位于伊犁谷地,均属于温带大陆性相对较为湿润的气候区。土壤均为山地黑棕色野苹果林土。植被的垂直分布比较明显,在大莫合沟和小莫合沟调查区,沟谷地带(海拔<1 300 m)主要乔木树种有新疆野苹果、野杏(Armeniaca vulgaris)和欧洲山杨(Populus tremula),偶见天山桦(Betula tianschanica),林下灌木稀少,偶有忍冬(Lonicera caerulea)和黑果小檗(Berberisheteropola),草本层以禾本科为主,盖度50% ~ 90%,主要伴生种有蒿子(Artemisia vulgaris)、草原糙苏(Phlomis pratensis)和焮麻(Urtica cannabina)等; 中海拔地带(海拔1 300 ~ 1 600 m)乔木层树种主要为新疆野苹果,灌木稀少,偶见有野蔷薇(Rosa spp.),林下草本层优势种为短距凤仙花(Impatiensbrachycentra)或羊角芹(Aegopadium podagraria),盖度在60%以上; 高海拔地带(海拔>1 600 m)乔木层的优势种为新疆野苹果,伴生种为天山云杉(Picea schrenkiana var.tianschanica),灌木稀少,草本层仍以禾本科为主,优势种有羊茅(Festuca ovena)、早熟禾(Poa annua)和红花三叶草(Trifolium pratense)等。各调查区自然概况见表 1。
|
|
分别在天山西部伊犁州的大莫合沟、小莫合沟和交吾托海沟海拔1 200 ~ 1 700 m新疆野苹果分布区各设置3条垂直调查样带,共9条。在每一样带内沿海拔梯度每50 m间隔设置1块20 m × 20 m的样地,由于山体条件不同(有些地段不适宜调查)每条样带内设置样地数不同,为8 ~ 11块。由于野苹果林的平均枝下高在1 m左右,故野外样方调查时选择新疆野苹果林木的地径来代替胸径,其他树种均调查胸径。调查内容包括: 1)种群特征对样地内所有乔木进行每木调查并记录种名、地径(或胸径)、树高、枝下高和冠幅; 并沿样地的2条对角线采用样点法测定样地林分的郁闭度; 2)林分环境因子利用GPS在样地中心位置记录样地的地理坐标和海拔高度,用罗盘仪实测样地坡度,同时记录样地所在坡向和坡位; 3)人为干扰状况在每一样地内和周围记录打草、放牧、居民点、农田、河流等人类活动或自然环境状况; 4)病虫害在每一样地内,记录病虫害为害的新疆野苹果株数。
2.2 大小结构划分本研究采用地径(D)和树高(H) 2个指标进行种群结构分析。在每个地点,根据新疆野苹果林样方调查数据,按上限排外法,将径级结构共划分13级: Ⅰ(1 cm≤D<5 cm),Ⅱ(5 cm≤D<10 cm),Ⅲ(10 cm ≤ D<15 cm),Ⅳ (15 cm ≤ D<20 cm),Ⅴ(20 cm≤ D<25 cm),Ⅵ (25 cm ≤ D<30 cm),Ⅶ(30 cm≤ D<35 cm),Ⅷ (35 cm ≤ D<40 cm),Ⅸ(40 cm≤ D<45 cm),Ⅹ (45 cm ≤ D<50 cm),Ⅺ(50 cm≤D<55 cm),Ⅻ (55 cm≤ D<60 cm)和ⅩⅢ(D≥60 cm); 将树高分为9级: Ⅰ(H<2 m),Ⅱ(2 m≤H<4 m),Ⅲ (4 m≤H<6 m),Ⅳ (6 m≤H<8 m),Ⅴ (8 m ≤ H<10 m),Ⅵ (10 m ≤ H<12 m),Ⅶ(12 m≤H<14 m),Ⅷ(14 m≤H<16 m)和Ⅸ(H≥16 m)。
2.3 数据处理利用SPSS 13.0进行统计分析和检验。
3 结果与分析 3.1 种群分布特征调查区新疆野苹果林主要分布在天山山体的阴坡或半阴坡。在景观上呈大小不同的斑块镶嵌,形成以新疆野苹果为优势种的单优群落。新疆野苹果大都分布在海拔1 200 ~ 1 700 m范围内。在海拔1 300 ~ 1 600 m处,新疆野苹果林以纯林形式分布,其面积占全部调查林分面积的71%。在其他海拔处的新疆野苹果林分伴生有少量的野杏、野生樱桃李(Prunus cerasifera)、天山云杉、天山桦、欧洲山杨和准噶尔山楂(Crataegus songorica)等树种:在较高海拔处新疆野苹果常与天山云杉伴生; 在海拔1300 m以下新疆野苹果多与野杏、天山桦和欧洲山杨等伴生。
3.2 种群径级和树高结构图 1表明,3个典型分布区新疆野苹果种群的个体数量随着径级的变化趋势一致,均呈单峰型分布:在Ⅱ~ Ⅵ径级分布的株数比例较多,小径级幼树和大径级树木所占比例较少。由于树木径级和龄级间存在正相关性,新疆野苹果种群的中龄级个体所占比例较大,种群配置不太均匀。大莫合沟种群径级结构相对完整,Ⅰ径级(D<5 cm)个体数量仅占该种群数量的2 %,Ⅱ~ Ⅵ径级占73.8 %,Ⅶ ~ Ⅸ径级占20.9 %,大于Ⅹ级的仅占3.3 %。小莫合沟内新疆野苹果种群的径级结构不完整,未见Ⅰ径级的个体分布,Ⅱ~ Ⅵ径级的个体数量占总体的67.4%,Ⅶ ~ Ⅸ径级占18.6%,大于Ⅹ级的占14.0 %。交吾托海沟内新疆野苹果种群的径级结构也不完整,Ⅰ和Ⅻ径级的个体均未出现,Ⅱ~ Ⅵ径级的个体数量占70.7 %,Ⅶ ~ Ⅸ径级占20.0 %,大于Ⅹ级的占9.3 %。小莫合沟和交吾托海沟种群中大于Ⅹ级的个体数量均大于大莫合沟。大莫合沟、小莫合沟和交吾托海沟3个新疆野苹果种群的平均地径分别为(21.10 ± 0.97),(27.42 ± 2.04)和(24.82 ± 1.19)cm,大莫合沟种群的平均地径显著P<0.05)低于小莫合沟和交吾托海沟,但小莫合沟和交吾托海沟的平均地径间差异不显著(P>0.05)。

|
图 1 种群径级和树高结构 Figure 1 Diameter and tree height class of Malus sieversii populations in three typical locations |
3个典型分布区新疆野苹果种群的树高结构均呈单峰型(图 1)。大莫合沟新疆野苹果种群的树高结构完整,而小莫合沟和交吾托海沟的种群中Ⅶ级以上高度的个体数量很少或没有出现。大莫合沟和小莫合沟新疆野苹果的平均树高分别为(7.05 ± 0.30)和(6.56 ± 0.29) m,2者间无显著差异性(P>0.05),但分别与交吾托海沟的平均树高(7.95 ± 0.18) m存在显著差异(P<0.05)。
相关分析结果表明,3个典型分布区树高和地径间均具有显著正相关性(P<0.05)。新疆野苹果3个种群平均树高随不同径级的分布均呈单峰型(图 2)。在相同的径级下,不同种群的新疆野苹果个体高度间差异显著(P<0.05)。

|
图 2 3个新疆野苹果种群不同径级下的平均树高 Figure 2 Mean tree height of three M.sieversii population in different diameter class |
在1 200 ~ 1 700 m海拔范围内,以新疆野苹果为单优种群的林分因子如树高、地径、枝下高、冠幅、郁闭度及林分密度随海拔的变化见图 3。在海拔1 200 ~ 1 700 m处,3个典型分布区以新疆野苹果为单优种群林分的地径和枝下高均随海拔的升高而降低。大莫合沟和交吾托海沟树高和冠幅随海拔升高而下降,小莫合沟树高和冠幅呈单峰型变化趋势。调查区内所有以新疆野苹果为单优种群的林分的平均郁闭度比较低,为0.48 ± 0.03,其中大莫合沟内的林分郁闭度仅为(0.38 ± 0.04),与小莫合沟(0.55 ± 0.09)和交吾托海沟(0.51 ± 0.08)的差异显著(P<0.05)。小莫合沟和交吾托海沟林分郁闭度随海拔的变化可认为是单峰型,低海拔和高海拔地段的郁闭度均低于中海拔; 大莫合沟内林分郁闭度随海拔的变化与上述2个样地相反,即中海拔地区的郁闭度分别较低海拔和高海拔低。调查区样地内林分密度随海拔的变化趋势与郁闭度与海拔间的变化规律一致。小莫合沟的平均林分密度为(502.3 ± 151.1)株·hm-2,是3个调查区中最高的,大莫合沟和交吾托海沟的林分密度值接近,分别为(319.2 ± 44.1)和(325.0 ± 91.1)株·hm-2; 对比林分密度和林分郁闭度随海拔梯度的变化趋势可以看出,大莫合沟内新疆野苹果在海拔1 500 m附近林分密度较小,郁闭度也相对较低; 小莫合沟的新疆野苹果林分在海拔1 450 ~ 1 500 m地段的密度达到最大,林分郁闭度也较高; 交吾托海沟在整个调查范围内的林分密度较前2个样地相对均匀,在海拔1 400 ~ 1 550 m处林分密度较高,而林分郁闭度在海拔1 300 m处最大。

|
图 3 新疆野苹果林分因子随海拔梯度的变化 Figure 3 Elevational distribution of stand factors dominated by M.sieversill forest |
大莫合沟新疆野苹果种群结构较小莫合沟和交吾托海沟完整,但林分密度和林分郁闭度均低于小莫合沟和交吾托海沟。3个分布区新疆野苹果种群的配置不均匀,更新幼苗和成熟甚至过熟林木数量极少,种群处于衰退状态。多年来,开发利用野果林资源引起了生态系统的衰退和演变,诸如当地兴建的野生果品加工企业在果实近成熟期大量采收果实,影响了新疆野苹果的生长发育、种子成熟及种群更新,是造成野苹果种群衰退的原因之一。林牧矛盾问题是导致新疆野苹果种群内幼苗稀缺、种群更新困难的又一原因:当地哈萨克族等牧民在野果林分布区过度放牧,林下植被成为家畜饲料的主要来源,大量的新疆野苹果根蘖苗和实生幼苗连同周围的杂草均被当作牧草刈割,结果导致新疆野苹果固有的繁育体系遭到破坏。在相同径级下3个新疆野苹果种群的平均树高间差异显著。在大径级个体中,其高生长速度减小,另外有树冠上部部分枝条死亡,所以其高度反而比中间径级小。
为保存珍贵的野生种质资源,改善新疆野苹果林分生长环境,维持其结构和功能稳定,需要认识到引发新疆野苹果林濒危的原因,并采用合理的方法。前人在相同地点的研究结果显示新疆野苹果分布在海拔1 100 ~ 1 800 m,在个别谷地未充分开垦前可低至1 000 m左右,高可达2 023 m (张新时,1973; 苏馨花等,2007)。本次调查结果显示,新疆野苹果的分布范围为海拔1 180 ~ 1 700 m,较前人的结果有所减小,分布下限上升。一方面垦殖和超载放牧致使大面积野果林被破坏,另一方面,近几年苹果小吉丁虫危害使大量的新疆野苹果死亡,这些都给新疆野苹果的生存造成了一定的压力,使得新疆野苹果不断地向较高海拔地区的山沟退缩。为了保存优良种源,可以在原有保护区范围的基础上扩大就地保育的面积,同时开展异地繁殖和保育研究。为了减少小吉丁虫对新疆野苹果的危害,在主要分布区内应封育或设置隔离带,并在4月中旬到5月初,用药剂(如80%敌敌畏乳剂)涂抹于枝条的病斑处,以杀死幼虫和抑制小吉丁虫卵的分化和病害的蔓延。山地森林的纬度、经度和海拔是影响物种分布格局的三大地理因素,前2种决定着物种在不同区域内的分布,而后者往往是同一区域内物种分布格局的决定因素(Wimberly et al., 2001; Zhao et al., 2005)。3个分布区新疆野苹果种群除物种组成有差异外,在整个分布区内,随着海拔的升高,新疆野苹果种群的树高随海拔的升高呈抛物线型分布,地径和枝下高随海拔的升高而降低,冠幅随海拔升高而增加。而且中低海拔和高海拔地段的林木个体数量均低于中海拔的。因此中间海拔范围是野苹果保育的重点区域,是建立野苹果保护区的核心地带。
新疆野苹果属于国家二级濒危保护树种。前人对新疆野苹果属于濒危树种的界定是基于其种质资源的分布范围、树木的生存数量和质量之上。而本研究从种群生态学角度,利用标准地调查数据分析3个典型分布区新疆野苹果种群结构,为进一步认识其濒危机理提供了理论基础。
崔大方, 廖文波, 羊海军, 等. 2006. 中国伊犁天山野果林区系表征地理成分及区系发生的研究[J]. 林业科学研究, 19(5): 555-560. |
冯涛, 张红, 陈学森, 等. 2006. 新疆野苹果果实形态与矿质元素含量多样性以及特异性状单株[J]. 植物遗传资源学报, 7(3): 270-276. |
傅立国. 1992. 中国植物红皮书-稀有濒危植物[M]. 北京: 科学出版社.
|
侯博, 许正. 2005. 天山伊犁谷地珍稀野林资源研究[J]. 西北植物学报, 25(11): 2266-2271. DOI:10.3321/j.issn:1000-4025.2005.11.022 |
姜彦成, 党荣理. 2002. 植物资源学[M]. 乌鲁木齐: 新疆人民出版社.
|
林培钧, 崔乃然. 2002. 天山野果林资源———伊犁野果林综合研究[M]. 北京: 中国林业出版社.
|
刘爱华, 阿里木, 徐毅, 等. 2007. 新疆野果林苹果小吉丁幼虫空间分布型研究[J]. 西北林学院学报, 22(6): 92-94. |
刘立诚, 排祖拉, 徐华君. 1999. 新疆伊犁和塔城地区野果林下土壤特性及系统分类研究[J]. 土壤通报, 30(4): 153-156. |
佘定域. 1995. 伊犁野果林土壤的形成条件、成土过程及其特性[J]. 土壤通报, 24(6): 260-263. |
苏馨花, 何琼, 司洪章. 2007. 伊犁河流域珍贵野果林资源的保护与利用[J]. 中国林业, (1): 32-33. |
王磊, 许正, 晁海, 等. 2006. 新疆霍城县大、小西沟野果林种子植物组成及资源[J]. 干旱区研究, 23(3): 446-452. |
阎国荣, 许正. 2001. 天山野生果树主要病害及其分布[J]. 干旱区研究, 18(2): 47-49. |
羊海军, 崔大方, 许正, 等. 2003. 中国天山野果林种子植物组成及资源状况分析[J]. 植物资源与环境学报, 12(2): 39-45. |
张新时. 1973. 伊犁野果林的生态地理特征和群落学问题[J]. 植物学报, 15(2): 239-246. |
Christopher M R, Gayle M V, Ann A R, et al. 2009. Genetic diversity and population structure in Malus sieversii, a wild progenitor species of domesticated apple[J]. Tree Genetics & Genomes, 5(2): 339-347. |
Condit R, Ashton P S, Baker P, et al. 2000. Spatial patterns in the distribution of tropical tree species[J]. Science, 288(5470): 1414-1418. DOI:10.1126/science.288.5470.1414 |
Crawly M J. 1986. Plant Ecology[M]. London: Blackwel Scientific Publications: 97-185.
|
Hou J H, Mi X C, Liu C R, et al. 2004. Spatial patterns and associations in a Quercus-Betula forest in northern China[J]. Journal of Vegetation Science, 15(3): 407-414. |
Luby J J, Alspach P A, Bus V G M, et al. 2002. Field resistance to fire blight in a diverse apple (Malus sp.) germplasm collection[J]. Journal of the American Society for Horticultural Science, 127(2): 245-253. |
Münzbergová Z. 2004. Effect of spatial scale on factors limiting species distributions in dry grassland fragments[J]. Journal of Ecology, 92(5): 854-867. DOI:10.1111/jec.2004.92.issue-5 |
Nagashima K, Yoshida S, Hosaka T. 2009. Patterns and factors in earlystage vegetation recovery at abandoned plantation clearcut sites in Oita, Japan: possible indicators for evaluating vegetation status[J]. Journal of Forest Research, 14(3): 135-146. DOI:10.1007/s10310-009-0118-8 |
Volk G M, Richards C M, Reilley A A, et al. 2005. Ex situ conservation of vegetativelypropagated species: development of a seed-based core collection for Malus sieversii[J]. Journal of the American Society for Horticultural Science, 130(2): 203-210. |
Wimberly M C, Spies T A. 2001. Influences of environment and disturbance on forest patterns in coastal oregon watersheds[J]. Ecology, 82(5): 1443-1459. DOI:10.1890/0012-9658(2001)082[1443:IOEADO]2.0.CO;2 |
Yan G, Hong L, Song W, et al. 2008. Genetic polymorphism of Malus sieversii populations in Xinjiang, China[J]. Genetic Resources and Crop Evolution, 55(1): 171-181. DOI:10.1007/s10722-007-9226-5 |
Zhang C, Chen X, He T, et al. 2007. Genetic structure of Malus sieversii population from Xinjiang, China, revealed by SSR markers[J]. Journal of Genetics and Genomics, 34(10): 947-955. DOI:10.1016/S1673-8527(07)60106-4 |
Zhao C M, Chen W L, Tian Z Q, et al. 2005. Altitudinal pattern of plants species diversity in Shennongjia mountains, central China[J]. Journal of Integrative Plant Biology, 47(12): 1431-1449. DOI:10.1111/jipb.2005.47.issue-12 |
Zhou Z Q, Li Y N. 2000. The RAPD evidence for the phylogenetic relationship of the closely related species of cultivated apple[J]. Genetic Resources and Crop Evolution, 47(4): 353-357. DOI:10.1023/A:1008740819941 |