 2010, Vol. 46
2010, Vol. 46文章信息
- 曹福亮, 蔡金峰, 汪贵斌, 张往祥
- Cao Fuliang, Cai Jinfeng, Wang Guibin, Zhang Wangxiang
- 淹水胁迫对乌桕生长及光合作用的影响
- Effects of Waterlogging Stress on the Growth and Photosynthesis of Sapium sebiferum
- 林业科学, 2010, 46(10): 57-61.
- Scientia Silvae Sinicae, 2010, 46(10): 57-61.
-
文章历史
- 收稿日期:2009-05-18
- 修回日期:2009-11-16
-
作者相关文章


水淹是植物主要非生物胁迫之一, 它可以发生在江河漫滩、盐沼、沼泽等多个生态系统中(谭淑端等, 2009)。我国长江、黄河等流域由于生态环境的恶化, 洪涝灾害频繁发生, 造成巨大的经济损失。随着人类对湿地干扰的日益剧烈, 湿地植被严重退化, 动植物种类迅速减少, 生物多样性下降。因此在湖泊河流源头地区植树造林, 改善生态环境, 培育强抗涝性种植材料是解决我国涝渍土地利用和沿海、沿江地带洪涝灾害以及生物多样性下降等问题的根本途径。
乌桕(Sapium sebiferum)属大戟科(Euphorbiaceae)乌桕属(Sapium)植物, 是我国重要的经济树种。乌桕种子可榨取桕脂和桕油, 两者皆为重要的工业用油料; 乌桕木材质地坚、韧、细致, 用途广泛; 叶、根、皮均可人药, 其根皮叶及木材的浸出液可做杀虫剂(刘东泽等, 2003); 另外, 由于乌桕树季相变化丰富, 园林用途也十分广泛(张小红, 2006)。
乌桕喜湿润气候环境, 且能忍受较长时期的浸泡(张克迪等, 1994), 国外有学者对乌桕在淹水胁迫及盐淹胁迫下生长状况做了初步的研究(Conner et al., 1997), 但国内外对乌桕耐涝机理报道较少。因此, 本试验研究乌桕在淹水胁迫下生长和光合特征的变化情况, 为乌桕耐涝的生理机制研究提供参考, 并为乌桕在园林绿化以及栽培中充分发挥作用提供一定的理论基础。
1 材料与方法 1.1 试验材料及试验设计试验材料为1年生乌桕实生苗, 分别来自浙江兰溪和福建漳浦。
2007年2月中旬, 挑选生长一致的苗木, 进行盆栽(直径30 cm, 高30 cm), 每盆2棵, 放置于南京林业大学森林培育学科技示范园内, 为保证苗木的相对一致性, 定植后进行平茬处理, 苗木生长期间正常浇水和维护管理。2007年6月24日每种源挑选40盆生长一致的苗木, 分为4组, 进行淹水处理, 将盆放在水槽里, 水槽根据设计注水, 4个处理水平分别为:A水平为对照处理, 土壤含水量为田间持水量的75%左右; B水平为轻度淹水胁迫, 水面在土壤表面以下15 cm, 模拟地下水位过高; C水平模拟自然状态下水渍危害, 土壤含水量处于完全饱和状态; D水平模拟涝害, 土壤含水量过饱和, 水面高于土面4 cm左右。
1.2 测定方法 1.2.1 苗高、地径的测定苗高用卷尺测量(精确到0.1 cm), 地径测定部位为高于土面6 cm处, 每株用游标卡尺交叉测定2次(精确到0.02 mm), 取平均值。
1.2.2 生物量的测定淹水处理前和结束时各测1次, 分根、茎、叶3部分分别称重, 在80 ℃烘箱内烘干至衡重, 计算生物量及其分配。
1.2.3 光合作用的测定用PP-System公司的CIRAS-1型光合作用仪测定, 每种源每处理测定6片叶子。
2 结果与分析 2.1 淹水胁迫对不同种源乌桕生长的影响 2.1.1 淹水胁迫下乌桕的外观表现及根系生长状况浙江种源在B, C处理下, 叶片形态与对照基本一致, 在D处理中后期, 下部叶片变黄脱落, 上部叶片出现紫色斑点。福建种源在B水平处理下, 叶片形态与对照无显著差异, 在C, D处理下, 淹水至15 ~ 20天时, 生长几乎停滞, 下部叶片枯萎死亡, 至24天时, D处理全部死亡, 淹水45天时, C处理下所有植株死亡。
B处理下, 浙江种源根系生长与对照基本一致, 至淹水20天, C处理下茎基部产生白色肥大皮孔, 茎与土壤接触部位产生不定根, 浸没在水中的根系侧根数量减少, 颜色变黑, 同时有一些粗的不定根产生; D处理下, 在淹水部位的茎上长出不定根并产生颜色发白的肥大皮孔; 随淹水时间的延长, C, D 2处理下不定根的数量进一步增多, 但初生非木质化的根几乎全部发黑死亡。福建种源在整个淹水过程中, 没有产生不定根和肥大的皮孔。根据淹水胁迫下2种源乌桕外观表现及根系生长状况可知:浙江种源乌桕的耐涝性较福建种源强。
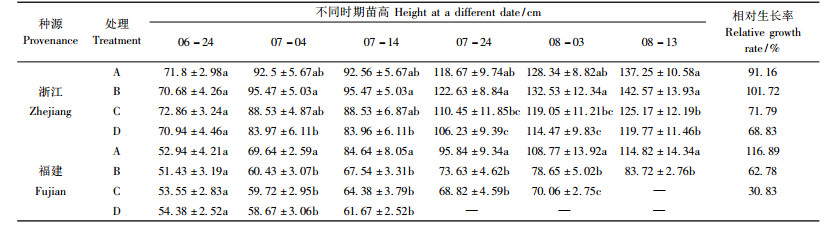
2.1.2 淹水胁迫对乌桕苗高的影响淹水胁迫下2种源乌桕苗高生长差异显著(表 1)。与对照相比, B处理促进浙江种源苗高的生长, 认为该种源乌桕可以适应地下水位较高的环境且生长良好; C, D处理早期, 高生长并没有受到明显的影响, 直到淹水20天后, 生长速度才明显减慢, 且随涝渍胁迫强度的加强, 苗高所受到的影响也越明显。福建种源各处理下高生长均受到明显抑制。
|
|
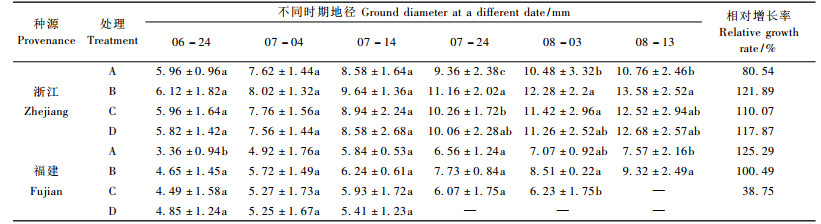
B, C, D处理均促进浙江种源地径的生长, 而以B处理下增长率最大(表 2)。C处理对福建种源地径增长的抑制作用非常显著, 至淹水30天时几乎停止生长, 而在淹水的末期甚至有一个降低的过程, 可能是由于较长时间的淹水处理, 韧皮部和皮层开始腐烂导致。
|
|
|
|
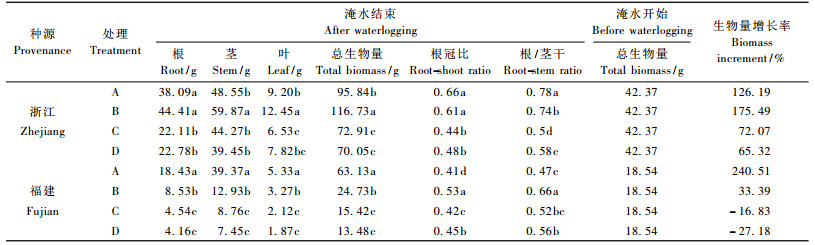
不同淹水处理对乌桕生物量增长造成显著影响(表 3)。与处理前相比, 浙江种源在B处理下总生物量较对照明显提高, C, D处理则抑制生物量积累。福建种源在淹水胁迫下生物量均明显低于对照, 在涝渍胁迫下生物量增长率出现负增长, 可能原因是绝大部分的初生根系已经腐烂, 且苗木落叶现象严重。
对2种源乌桕根冠比进行比较, 浙江种源在B处理下与对照差异不显著, 说明轻度的淹水并不对其造成影响; 福建种源在B处理下的根冠比高于对照, 分析可知:在轻度水分胁迫下, 其根、冠与对照相比相对增长率分别为-53.71%和-63.76%, 即根系减少的幅度比较小, 所以出现根冠比反而大于对照的现象。
2.2 淹水胁迫对乌桕净光合作用的影响 2.2.1 淹水胁迫对乌桕净光合速率的影响淹水10天时, 与对照相比, 2种源在淹水胁迫下净光合速率明显下降, 且随胁迫程度的加重, 降低的幅度增大(图 1)。浙江种源在3个水平的淹水处理下, 净光合速率分别比对照下降37%, 53%和59%, 而福建种源下降幅度分别为32%, 49%和79%。淹水30天时, 浙江种源在B处理下的净光合速率比对照高, 在C, D处理下与对照相比降低30%和34%, 与淹水10天时相比, 降低的幅度有所回升; 而福建种源的变化趋势与淹水10天时的趋势相同, 且下降的幅度有增加的趋势, 在C处理下净光合速率仅为2.52 μmol·m2 s, 与对照相比, 下降87%。淹水结束时, 浙江种源在B处理下高于对照22%, 在C, D处理下净光合速率则比对照下降37%和42%, 与淹水30天时相比下降的幅度有所加大; 福建种源在B, C, D处理下净光合速率显著下降。

|
图 1 淹水胁迫对净光合速率的影响 Figure 1 The effect of water logging stress on net photosynthesis rate |
淹水胁迫下, 乌桕叶片气孔导度的变化与净光合速率的变化相似, 即对照和B处理比C, D处理的高, 但在不同的测定时间又表现出一定的差异(图 2)。

|
图 2 淹水胁迫对气孔导度的影响 Figure 2 The effect of water logging stress on stomatal conductance |
淹水10天时, 与对照相比, 浙江种源在各淹水胁迫下气孔导度分别下降24%, 76%和75%;福建种源下降的幅度分别为63%, 58%和81%。淹水30天时, 浙江种源在B处理下与对照差异不显著, 在C, D处理下分别比对照下降63%和54%, 下降幅度较10天时有所回升; 福建种源气孔导度则随淹水时间延长逐渐下降, 在B, C处理下分别比对照低57%和94%。淹水结束时, B处理下浙江种源比对照高46%, C, D处理比对照下降59%和55%;福建种源在B处理下气孔导度仅为76 mmol·m-2 s-1, 与对照相比下降85%, 而涝渍胁迫下的植株均已死亡。
2.2.3 淹水胁迫对乌桕胞间CO2浓度的影响在淹水胁迫下, 浙江种源叶片细胞间CO2浓度与气孔导度的变化趋势一致, 即在淹水过程中, C, D处理下叶片细胞间CO2浓度要显著地低于对照和B处理(图 3)。福建种源细胞间CO2浓度呈现出随胁迫程度加重逐渐升高的趋势, 并不随着气孔导度的下降而下降。

|
图 3 淹水胁迫对胞间CO2浓度的影响 Figure 3 The effect of water logging stress on internal CO2 concentration |
淹水过程中, 浙江种源出现肥大皮孔和不定根, 这是耐淹木本植物对淹水的标志性形态反应(靖元孝等, 2000), 肥大皮孔、通气组织和不定根, 可以提高露于空气中吸收O2组织的总面积(Kozlowski, 1997)。福建种源没有形成不定根和肥大皮孔, 这可能是其对淹水敏感的原因之一。
研究表明:涝渍胁迫影响乌桕的主干生长, 使其苗高生长降低。曹福亮等(1993)认为淹水造成苗木高生长降低的直接影响是阻碍节间的生长, 间接作用是通过阻止叶原基分化、叶片生长以及诱导叶衰老、脱落等。淹水处理下, 浙江种源地径增长比对照大, 可能是由于形成肥大的皮孔以及在茎基部形成通气组织所致。
涝渍处理显著地抑制乌桕各器官的生长, 阻碍生物量的增加, 这与唐罗忠等(1998)对2种杨树(Populus)无性系的研究结果一致。B处理则提高了浙江种源的生物量, 与侯嫦英(2003)和杨静(2007)对落羽杉(Taxodium distichum)的研究结果一致, 认为浙江种源对水淹不敏感, 为喜湿树种, 湿度的水分胁迫可以促进其生长。
曹福亮等(1993)研究发现:美洲黑杨无性系在淹水处理后, 根冠比增大; 唐罗忠等(1998)研究发现, 杨树苗木根冠比均随着水分胁迫强度的增加而提高。本试验研究表明:2种源在涝渍胁迫下根冠比与上述研究有所不同, 即均要显著地低于对照和B处理, 表明涝渍胁迫对地下部分生长的影响更严重。
3.2 淹水胁迫对乌桕光合作用的影响植物对土壤淹水缺氧最敏感的响应就是气孔关闭, 气孔导度下降, 叶片吸收CO2的能力降低, 从而降低胞间CO2浓度, 而光合酶的底物变少则直接导致净光合速率的降低(Farquhar et al., 1982)。
本试验研究发现:涝渍胁迫下, 2种源乌桕叶片净光合速率和气孔导度与对照相比, 均有不同程度的降低, 这与McLeod等(1996)对包括乌桕在内的4种造林树种在盐淹胁迫下的研究结果一致, 与烟草(Nicotiana tobacum)(曾淑华等, 2005)、鹅掌楸(Liriodendron chinense)(张晓平, 2004; 潘向艳, 2006)、枫杨(Pterocarya stenoptera)和栓皮栎(Quercus variabilis)(衣英华等, 2006)、网纹甜瓜(Cucumis melo)(刘义玲等, 2009)等的研究结果基本一致; 轻度的淹水胁迫则促进浙江种源净光合速率的增加, 与淹水下落羽杉(杨静, 2007)的研究结论一致。
相关分析表明:浙江种源在整个涝渍处理过程中, 胞间CO2浓度与气孔导度的相关系数为0.868, 所以认为浙江种源净光合速率的下降主要是由气孔限制所致; 而福建种源在淹水处理的后期, 胞间CO2浓度并不随着气孔导度降低而下降, 反而有所升高, 按照Farquhar等(1982)的观点, 即只有当胞间CO2浓度的降低和气孔限制值增大时, 才可以做出光合速率的下降是由气孔导度的下降所引起的结论, 推测福建种源在淹水后期光合速率的下降主要是由非气孔限制所致。
本试验还发现:在涝渍胁迫下, 浙江种源净光合速率和气孔导度并不是随着淹水时间的持续而逐渐的下降, 而是在淹水胁迫的中期均有不同程度的回升, 这与水翁(Cleistocalyx operculatus)(靖元孝等, 2000)和枫杨(衣英华等, 2006)的研究结果相似, 可能是由于浙江种源乌桕在淹水过程中产生了不定根以及与水接触的茎基部形成肥大的皮孔, 有效弥补地下根系的低氧状态, 从而各项生理指标有一定幅度的回升, 以适应淹水环境。
曹福亮, 罗伯特·法门. 1993. 人工淹水胁迫处理对美洲黑杨无性系苗形态和生长的影响[J]. 南京林业大学学报, 17: 39-44. |
侯嫦英. 2003. 水分胁迫对青檀等树种生长及生理特性的影响. 南京林业大学博士学位论文.
|
靖元孝, 陈兆平, 程惠青, 等. 2000. 淹水时水翁幼苗光合特性与不定根的关系[J]. 热带亚热带植物学报, 8(4): 361-364. |
刘东泽, 赵杰, 刘志诚, 等. 2003. 20种植物提取物对几种昆虫的拒食作用[J]. 农药学学报, 5(3): 89-91. |
刘义玲, 李天来, 孙周平, 等. 2009. 根际低氧胁迫对网纹甜瓜光合作用、产量和品质的影响[J]. 园艺学报, 36(10): 1465-1472. DOI:10.3321/j.issn:0513-353X.2009.10.010 |
潘向艳. 2006. 杂交鹅掌楸不同无性系对淹水胁迫的反应. 南京林业大学博士学位论文.
|
谭淑端, 朱明勇, 张克荣, 等. 2009. 植物对水淹胁迫的响应与适应[J]. 生态学杂志, 28(9): 1871-1877. |
唐罗忠, 徐锡增, 方升佐. 1998. 土壤涝渍对杨树和柳树苗期生长及生理性状影响的研究[J]. 应用生态学报, 9(5): 471-474. |
杨静. 2007. 几种城市森林优良景观生态树的抗涝性研究. 南京林业大学博士学位论文.
|
衣英华, 樊大勇, 谢宗强, 等. 2006. 模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响[J]. 植物生态学报, 30(6): 960-968. |
曾淑华, 赵正雄, 覃鹏, 等. 2005. 淹水对转超氧化物歧化酶或过氧化物酶基因烟草某些生理生化指标的影响[J]. 植物生理学通讯, 41(5): 603-606. |
张克迪, 林一天. 1994. 中国乌桕[M]. 北京: 中国林业出版社.
|
张小红. 2006. 乌桕的园林应用与栽培[J]. 中国林业, (09): 41. |
张晓平. 2004. 不同种源鹅掌揪和杂种鹅掌揪对淹水胁迫的响应. 南京林业大学博士学位论文.
|
Conner W H, McLeod K W, McCarron J K. 1997. Flooding and salinity effects on growth and survival of four common forest wetland species[J]. Wetlands Ecology and Management, 5(2): 99-109. DOI:10.1023/A:1008251127131 |
Farquhar G D, Sharkey T D. 1982. Stomatal conductance and photosynthesis[J]. Ann Rev Plant Physiol, 33: 317-345. DOI:10.1146/annurev.pp.33.060182.001533 |
Kozlowski T T. 1997. Responses of woody plants to flooding and salinity[J]. Tree Physiology Monograph, 1: 1-29. |
McLeod K W, McCarran J K, Conner W H. 1996. Effect of flooding and salinity on photosynthesis and water relations of four Southeastern Coastal plain forest species[J]. Wetlands Ecology and Management, 4(1): 31-42. DOI:10.1007/BF01876133 |