 2010, Vol. 46
2010, Vol. 46文章信息
- 刘青华, 张蕊, 金国庆, 储德裕, 周志春
- Liu Qinghua, Zhang Rui, Jin Guoqing, Chu Deyu, Zhou Zhichun
- 马尾松年轮宽度和木材基本密度的种源变异及早期选择
- Variation of Ring Width and Wood Basic Density and Early Selection of Pinus massoniana Provenances
- 林业科学, 2010, 46(5): 49-54.
- Scientia Silvae Sinicae, 2010, 46(5): 49-54.
-
文章历史
- 收稿日期:2009-03-19
-
作者相关文章



2. 浙江省淳安县姥山林场 淳安 311700
2. Laoshan Forest Farm of Chun'an County, Zhejiang Province Chun'an 311700
种源试验和优良种源选择是林木遗传改良最基础的工作。世界上一些重要造林树种欧洲赤松(Pinus sylvestris)、挪威云杉(Picea abies)、火炬松(P. taeda)、杉木(Cunninghamia lanceolata)、马尾松(P. massoniana)、油松(P. tabulaeformis)和落叶松(Larix)等已开展了大规模的种源试验,揭示了生长、形态和材质等主要经济性状的种源和地理变异规律,为不同地区选出了适用的优质高产优良种源。然而不同种源生长发育特性存在显著差异,有些种源表现出早期速生丰产,另一些种源早期生长速率一般但中后期生长较快。又如幼龄材管胞短,胞壁薄,密度变异大,成熟材细胞细长,胞壁厚,密度较稳定(Zobel et al., 1989),因此种源生长和材性变异存在着明显的年龄效应。研究各种源生长和材性随年龄的变化规律,揭示种源生长和材性的年龄效应,可降低在幼龄期对各性状早期选择而产生的偏差。
林木不同基因型(种源、家系)生长随树木发育的变化模式有很多相关研究报道,如Isik等(1995)发现挪威云杉无性系有早期生长慢后期加快型及早期生长快后期转慢型等多种类型; 秦国峰(2003)基于对49个马尾松种源材积生长量的生长进程研究,将种源分为快、慢、先快后慢和先慢后快4种类型。木材基本密度随年龄的径向变异模式因树种而异,但普遍认为硬松类树种的木材密度由髓心向外迅速增加,然后逐渐趋于平稳(Zobel et al., 1989)。鉴于幼龄材与成熟材材性的显著差异,降低幼龄材比例和提高幼龄材密度可以提高木材的均匀性(Zobel et al., 1989; Nicholls et al., 1980)。Loo等(1985)和Gapare等(2006)分别研究了火炬松和辐射松(Pinus radiata), 认为木材密度由幼龄材向成熟材过渡年龄分别在11和7.5年左右, 过渡年龄受中等强度的遗传控制,以期选择幼龄期短、幼龄材密度大、材性均匀的基因型。但已有报道以幼龄阶段或未达到主伐年龄时的研究结果为主。本文利用设置在浙江省淳安县姥山林场24年生49个产地的马尾松种源试验林,在马尾松生长、形质和木材基本密度种源变异与种源区划研究的基础上(刘青华等, 2009),进一步研究年轮宽度和木材基本密度的种源变异及年龄效应,阐明不同种源区种源随树龄的增加年轮宽度和基本密度的径向变异规律及木材密度径向均匀性的差异,揭示不同年轮段年轮宽度和木材基本密度的遗传相关和早晚期相关,从而为早期选择速生、材性均匀性高的马尾松种源提供重要理论依据。
1 材料与方法 1.1 材料来源试验材料取自设置在浙江省淳安县姥山林场的24年生马尾松种源试验林,来自14个省区49个产地的种源参试。该种源试验林位于浙江省千岛湖东南湖区,自然概况、试验设计及育林方式、措施见刘青华等(2009)。
1.2 试验林调查和木材基本密度测定由于重复Ⅰ和Ⅳ保存不完整,本文只利用其他6个重复的材料。2008年4月在每重复的试验小区中选择2株生长最佳植株,在测其树高、胸径、通直度等生长和形质的同时(刘青华等, 2009),在植株胸高处上坡方位用6 mm直径的生长锥钻取一髓心至树皮的完整无疵木芯。对所取木芯自髓心向外,每5个年轮切成一段,测其宽度(Wi),并用最大含水量法测每年轮段的基本密度(Di)(Smith, 1954)。
1.3 数据分析利用各年轮段宽度测定值估算其所代表的年轮段面积[Ai= π(∑li)2-π(∑li-1)2,li为第i段木芯的长度],进而计算1~5, 6~10, 11~15, 16~20, 21~22轮, 及幼龄材(1~10轮)和成熟材(16~22轮)木芯的加权平均基本密度Db=∑AiDi/ ∑Ai。以小区单株测定值为单元,采用SAS/STAT软件中的GLM程序进行性状方差分析,以验证重复、种源和重复×种源互作效应。性状表型/遗传相关分析时以小区平均值为单元,采用DPS数据分析软件处理。通过比较,选用y=ax3+bx2+cx+d三次多项式模型来拟合马尾松种源径向生长和木材基本密度与年龄的关系,以揭示其年龄效应,其中y为各年轮段的平均年轮宽度或各年轮段的木材基本密度,x为各年轮段距髓心的平均年轮数,a、b、c和d为回归常数。
2 结果与分析 2.1 不同年龄段年轮宽度和木材基本密度的种源差异表 1方差分析结果显示,马尾松1~5, 6~10, 11~15, 16~20, 21~22轮的年轮宽度皆存在着极显著的种源差异,各年轮段年轮宽度的变异系数都在20%以上,意味着在从幼林到主伐期间的不同生长发育阶段马尾松49个产地间的径向生长皆存在较大的差异,径向生长的种源效应贯穿整个树木生长发育阶段,由此可见种源选择对于提高马尾松生产力的重要性。从6~10轮向11~15轮和16~20轮年轮宽度的种源变异系数逐渐增大,其中以16~20轮年轮宽度的种源变异系数最大,达31.23%。其原因除与种源间本身的遗传差异有关外,还与因试验小区较小导致种源间生长竞争剧烈进而人为地加大种源间差异有关(周志春等, 1993)。
|
|
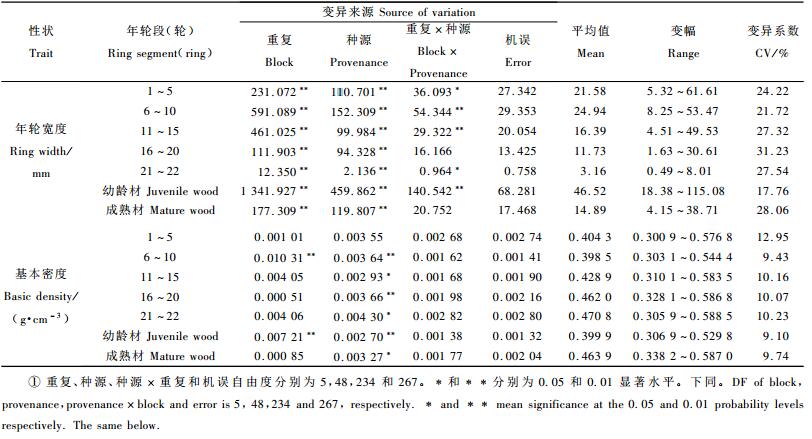
近髓部1~5轮的木材基本密度种源效应较小(表 1),进一步证实了周志春等(1990)的研究结果,而其他年轮段木材基本密度的种源效应皆达到显著或极显著水平。虽然1~5轮年轮段木材基本密度的种源效应不显著,但其种源变异系数却最大,达12.95%,意味着仍具有较大的选择潜力。第6轮后各年轮段木材基本密度的种源变异系数稳定在10%左右,较少受年轮生长发育、试验重复及其与种源互作的影响。综上说明通过种源选择可实现木材基本密度的遗传改良。
表 1还列出了马尾松种源幼龄材和成熟材年轮宽度和基本密度的方差分析结果,发现其皆存在显著或极显著的种源差异。相对于成熟材,幼龄材年轮宽度的种源变异系数较小,仅为成熟材的63.29%,表明马尾松径向生长的种源效应在成熟期表现得尤为明显。幼龄材和成熟材基本密度的种源变异系数差异却不大。
2.2 不同种源年轮宽度和木材基本密度的径向变异模式根据前文对24年生49个产地的种源试验结果,将马尾松划分为南部、中部和北部3个种源区(刘青华等, 2009)。图 1利用三次多项式分别3个种源区拟合了种源各年轮段平均年轮宽度的径向变化曲线,其R2在0.955 3~0.990 2间,拟合度很高。从图 1可以看出,3个种源区的种源各年轮段年轮宽度的径向变异趋势相似,皆表现出先迅速增长,在6~10轮年轮宽度达到最大值,之后各年轮段宽度逐渐变窄,21~22轮平均年轮宽度仅为6~10轮的31.67%。在径向生长过程中,南部种源一直快于中部和北部种源,这种趋势在6~10轮段表现最为明显,随后差异不断缩小,在21~22轮段3个种源区种源的平均年轮宽度相差较小。此外,根据年轮宽度连年生长量和平均生长量的相交位置(图 2)可推测,3个种源区种源的径向生长在16~20轮皆可达到数量成熟,但到达时间的先后依次为南部种源区、中部种源区和北部种源区的种源。

|
图 1 3个种源区种源平均年轮宽度的径向变化曲线 Figure 1 Radial change of average ring width with cambial age for P. massoniana provenances from three provenance zones |

|
图 2 3个种源区种源的连年生长量和平均生长量 Figure 2 Curves of current annual and average increment for P. massoniana provenances from three provenance zones |
图 3为3个种源区种源各年轮段木材基本密度随年龄变化的径向变异趋势线,皆表现为随年龄增加木材基本密度先增加后逐渐变平稳的“S”型,与年轮宽度的径向变化趋势正好相反。南部种源1~5轮木材基本密度测定值偏高,是由于松脂等在靠近髓心部位大量沉积、抽提物含量较高之故。在6~10轮和11~15轮段来自南部区种源的木材基本密度明显低于中部和北部区的种源,但在16~20轮和21~22轮段其间的差异却较小,这一结果说明对南部区种源幼龄材基本密度的遗传改良显得尤为必要。

|
图 3 3个种源区种源木材基本密度的径向变化曲线 Figure 3 Radial change of average wood basic density with cambial age for P. massoniana provenances from three provenance zones |
材性改良的一个主要目的是提高木材性状的均匀性,尤其是径向的均匀性。Zobel等(1989)认为基本密度的不均匀是木材最主要的缺陷,它严重影响最终产品的品质和制浆造纸的成本。提高幼龄材密度可降低因短轮伐期经营而造成木材品质下降的负面影响(Rozenberg et al., 1997)。这里用幼龄材与成熟材木材基本密度的比值(I)来表示基本密度径向变异的均匀性。南部区种源的I值变化在0.77~0.88(图 4),种源变异系数为4.31%;相对于南部区种源,中部区种源I的平均值提高了20%,但种源间木材基本密度径向均匀性却较差,种源I值变化在0.75~0.92。北部区种源木材基本密度I的平均值最高(0.89),种源间变化较小(0.84~0.91)。由此可见,部分南部区种源的快速生长是以降低幼龄材密度为代价的,致使幼龄材和成熟材基本密度差异较大。但从图 4可看出,也有部分南部区种源木材基本密度的径向均匀性较高。以49个参试种源的I均值为阈值,从已确定的19个纸浆材优良种源(刘青华等,2009)中进一步选择出广西横县、恭城、忻城,广东乳源和福建南靖5个基本密度径向均匀性高的纸浆材优良种源。

|
图 4 3个种源区种源的I值分布图 Figure 4 `Ratio of basic density in juvenile wood to that in mature wood (I) for P. massoniana provenances from three provenance zones |
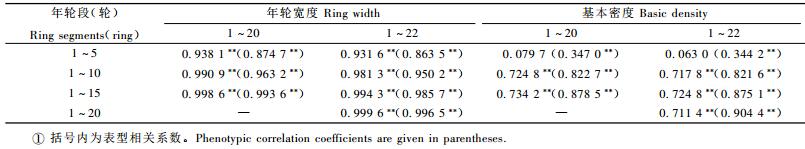
表 2性状相关分析表明,随着年龄增加和树木生长发育,马尾松种源年轮宽度和木材基本密度间的负相关性不管是在表型还是在遗传上都逐渐加强。1~5轮段年轮宽度和木材基本密度的表型和遗传均不相关,但在第6轮(约8年生)后,各年轮段年轮宽度和木材基本密度的负遗传相关都达到显著或极显著水平,选择径向生长快的种源会明显降低其木材基本密度。
|
|

表 3列出了1~5, 1~10, 1~15轮年轮宽度和木材基本密度与不同伐期(1~20, 1~22轮)时的早晚相关。从表中可看出1~5, 1~10, 1~15轮年轮宽度与1~20, 1~22轮的年轮宽度均呈极显著的表型和遗传相关,利用1~5轮段(约7年生)的年轮宽度即可对种源生长进行有效的早期选择。1~5轮的基本密度与1~20, 1~22轮的基本密度遗传相关性较小,但1~10, 1~15轮的基本密度与1~20, 1~22轮的基本密度的表型和遗传相关皆极显著(P < 0.01),表明木材基本密度早期选择的年龄晚于径向生长。因此进行高产、优质种源选择时,可在7年生(1~5轮)时先淘汰生长较差的种源,在12年时(1~10轮)再对种源进行生长和木材基本密度的选择。
|
|
在生长早期或幼龄材阶段,马尾松各种源的径生长相对较快,且较少受树木竞争影响,结果年轮宽度的种源变异较小。随着树木的生长发育,种源径生长量总体上有所减小,但由于部分种源在生长后期仍具有较大的径生长量,一些慢生种源径生长因植株间竞争而受抑,导致生长后期年轮宽度的种源效应明显增大,如幼龄材年轮宽度的种源变异系数仅为成熟材的63.29%,说明年轮宽度种源变异存在明显的年龄效应。通过比较发现,3个种源区种源年轮宽度、木材基本密度的径向变异模式相似,表现为年轮宽度随着形成层年龄的增加先增后降,年轮木材基本密度随着形成层年龄的增加先增后变平缓,呈“S”型的变化趋势,这类似于红松(Pinus koraiensis)(王慧梅等,2004)、加勒比松(P. caribaea)(文小明等,1996)和杉木(骆秀琴等,1999)等种源的研究结果,这也证实了Zobel等(1989)提出的硬松类树种木材密度从髓心向外先迅速增加,在接近树皮时逐渐变平稳的观点。
木材基本密度径向均匀性是衡量种源材性的重要指标,也是说明种源木材密度年龄效应的一个较好指标。研究表明,不同种源区种源木材密度径向变异均匀性存在着明显的差异。相对于北部和中部种源区种源,南部种源区种源在幼龄期其径生长明显较快,细胞壁薄、腔大、晚材比例低,结果其木材基本密度明显地低于中部和北部种源区的种源。而到了成熟材阶段,木材细胞变小、壁增厚、腔变窄,晚材比例增大,3个种源区种源的木材基本密度都变大而差异缩小,结果导致南部种源区种源木材密度的径向均匀性较低。然而在南部种源区种源中,仍存在一些速生、木材密度大、木材密度径向均匀性高的优良种源,其中广东乳源和福建南靖2种源表现最优。下一步研究组还将研究年轮木材密度组成及木材密度由幼龄材向成熟材过渡年龄的种源变异,以期为选择过渡年龄小、幼龄材密度大、径向均匀性高、年轮内差异小的速生优质种源提供重要理论依据。
生长速度对材性的影响较为复杂,涉及到遗传、环境和树龄等多个方面,也是一个颇具争议的内容。不同年轮段年轮宽度和木材基本密度的相关分析结果表明,除1~5轮段外,各年轮段年轮宽度和木材基本密度的负遗传相关都达到显著或极显著水平,进一步证实了已有研究结果(周志春等,1990; 1993),即选择生产力高的速生种源会明显降低其木材密度。在生长早期,种源木材基本密度均较小,种源间的差异不显著,但早期径生长的种源效应却较大,结果导致1~5轮段的年轮生长与木材基本密度相关性较小。早晚相关分析结果表明,各年龄段年轮宽度与主伐期时的年轮宽度高度相关,而木材基本密度约在10轮时(12年生)才能表现出与主伐期时木材密度的高度相关。这一结论可解释马尾松种源木材基本密度在幼龄期的地理变异模式(周志春等,1993; 徐立安等,1997)与已达主伐年龄时的地理变异模式(刘青华等,2009)的不同,即木材密度受年龄影响较大。在选择高产、优质种源时,可在7年生时先对生长较差的种源进行淘汰,在12年生时再对种源生长和木材基本密度进行联合选择。
刘青华, 金国庆, 张蕊, 等. 2009. 24年生马尾松生长、形质和木材基本密度种源变异与种源区划[J]. 林业科学, 45(10): 55-61. DOI:10.11707/j.1001-7488.20091010 |
骆秀琴, 管宁, 文小明, 等. 1999. 木材材性株内径向变异模式初探Ⅵ: 19个杉木种源木材密度径向变异模式的研究[J]. 林业科学, 35(6): 86-92. |
秦国峰. 2003. 马尾松地理种源[M]. 杭州: 浙江大学出版社: 74-79.
|
王慧梅, 夏德安, 王文杰. 2004. 红松种源材质性状研究[J]. 植物研究, 24(4): 495-498. |
文小明, 骆秀琴, 管宁, 等. 1996. 木材材性株内径向变异模式初探II.七个加勒比松种源木材密度径向变异模式的研究[J]. 林业科学, 32(5): 461-419. |
徐立安, 陈天华, 王章荣, 等. 1997. 马尾松种源子代材性变异与制浆造纸材优良种源选择[J]. 南京林业大学学报, 21(2): 1-6. |
周志春, 傅玉狮, 吴天林. 1993. 马尾松生长和材性的地理遗传变异及最优种源区的划定[J]. 林业科学研究, 6(5): 556-564. |
周志春, 金国庆, 秦国峰. 1990. 马尾松幼龄材密度、管胞长度的地理遗传变异及性状相关[J]. 林业科学研究, 3(4): 393-397. |
Gapare W J, Wu H X, Abarquez A. 2006. Genetic control of the time of transition from juvenile to mature wood in Pinus radiata D.Don[J]. Annals of Forest Science, 63(8): 871-878. DOI:10.1051/forest:2006070 |
Isik K, Kleinschmit J, Svolba J. 1995. Survival, growth trends and genetic gains in 17-year old Picea abies clones at seven test sites[J]. Silvae Genetica, 44(2/3): 116-128. |
Loo J A, Tauer C G, Mcnew R W. 1985. Genetic variation in the time of transition from juvenile to mature wood in loblolly pine (Pinus taeda L)[J]. Silvae Genetica, 34(1): 14-19. |
Nicholls J W, Morris J D, Pederick L A. 1980. Heritability estimates of density characteristics in juvenile Pinus radiata wood[J]. Silvae Genetica, 29(2): 54-61. |
Rozenberg P, Cahalan C. 1997. Spruce and wood quality: genetic aspects (a review)[J]. Silvae Genetica, 46(5): 270-279. |
Smith D M. 1954. Maximum moisture content method for determining specific gravity of small wood samples. US Forest Service, Forest Products Laboratory, Report No: 2014. https://www.fpl.fs.fed.us/documnts/fplr/fplr2014.pdf
|
Zobel B J, van Buijtenen J P. 1989. Wood variation: its causes and control[J]. Berlin: Springer-Verlag, 72: 72-131. |