 2010, Vol. 46
2010, Vol. 46文章信息
- 孟昭军, 周永泉, 严善春, 金虎, 胡晓
- Meng Zhaojun, Zhou Yongquan, Yan Shanchun, Jin Hu, Hu Xiao
- 外源茉莉酸类化合物对2种落叶松针叶内单宁含量的影响
- Effects of Exogenous Jasmonates on Tannin Content in Needles of Two Larch Species
- 林业科学, 2010, 46(3): 96-104.
- Scientia Silvae Sinicae, 2010, 46(3): 96-104.
-
文章历史
- 收稿日期:2009-02-04
-
作者相关文章



2. 黑龙江省大兴安岭阿木尔林业局 漠河 165300
2. Amur Forestry Bureau of Daxing'anling Mountains in Heilongjiang Province Mohe 165300
单宁是一类化学结构和生物活性复杂的酚类化合物,广泛存在于植物的皮、根、叶、果中,特别是针叶树中尤为普遍(Gebauer et al., 1998; Seigler,1998),质量分数常常超过10% (Heyworth et al., 1998)。单宁可以作为植物防止动物或微生物伤害的一种有效防护剂(Scalbert,1991),其收敛性即涩性可以减少植食者的取食,并与食物中的蛋白质、淀粉结合降低其营养价值,与植食者消化道内消化酶结合降低其消化能力,影响其对营养物质的吸收率,导致植食者营养不良(Muir et al., 1999); 其能延迟淡色库蚊(Culex pipiens pallens)幼虫、棉铃虫(Helicoverpa armigera) 1龄幼虫的生长发育(武予清等,2001; 张时妙等,2005); 100 mg·L-1单宁酸溶液可使五带淡色库蚊(Culex quinquef asciatus)幼虫完全死亡(Pospisil et al., 1981)。马尾松(Pinus massoniana)被马尾松毛虫(Dendrolimus punctatus)(戈峰等,1997)、松墨天牛(Monochamus alternatu)(李水清等,2007)取食或人为损伤后,针叶内单宁、总酚或黄酮类物质含量增加; 而赤松毛虫(D. spectabilis)为害的2年生油松(P. tabulaeformis)针叶(李镇宇等,1998; 1999) 及华北落叶松鞘蛾(Coleophora sinensis)取食的华北落叶松(Larix principis-rupprechtii)针叶(薛皎亮等,2000),它们针叶内单宁或生物碱含量均会增加。当环境中CO2的浓度提高时,欧洲赤松(P. sylvestris) (Räisänen et al., 2008)、火炬松(P. taeda) (Gebauer et al., 1998)针叶内单宁含量会升高。虫害取食或机械损伤还能诱导其他植物体内单宁含量增加,是植物诱导防御的重要物质(Peters et al., 2002; 程红等,2006)。应用外源茉莉酸(jasmonic acid,JA)或茉莉酮[(Z)-jasmone]也能诱导植物体内单宁含量的改变,影响昆虫的取食及其正常的生长发育,从而增强植物的自主化学防御(徐伟,2006; Cooper et al., 2008)。
落叶松(Larix spp.)是我国东北地区主要的造林树种,不同年份不同地区,其各类害虫均有不同程度的发生,受害严重。已有研究显示:用剪叶损伤、昆虫取食或模拟昆虫取食及1 mmol·L-1茉莉酸、茉莉酮、水杨酸(salicylic acid,SA)、水杨酸甲酯(methyl salicylate,MeSA)等不同因子分别处理兴安落叶松(L. gmelinii)后,其针叶内单宁含量明显增加,兴安落叶松的诱导抗性产生(徐伟,2006; 袁红娥等,2009)。本文在不同因子诱导兴安落叶松单宁含量变化研究的基础上,以兴安落叶松和长白落叶松(L. olgensis) 2种近缘植物为试验材料,通过施用不同浓度的外源茉莉酸类化合物(jasmonates,JAs),研究其对2种落叶松针叶内单宁含量变化诱导的作用,比较2种落叶松应激反应的差异,为不同种落叶松提供较为适宜的害虫治理对策。
1 材料与方法 1.1 供试落叶松于4月末将2年生的兴安落叶松、长白落叶松2种树苗各2 000株栽于帽儿山林场老山站育苗网棚内,自然光照条件下生长,保证落叶松苗有充足的水分,及时施肥、除草,待用。
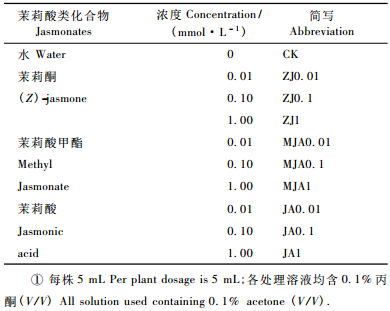
1.2 落叶松处理落叶松苗处理参照Heijari等(2005)和桂连友等(2005)的方法,并加以修改。于8月各选取长势相似的落叶松苗900株,随机分为10种处理,每种处理90株,用500 mL手动喷雾器将不同浓度外源茉莉酸化合物溶液均匀喷到各处理落叶松苗上,待用(表 1)。
|
|
分别在处理后第1,3,5,10天(兴安落叶松)和1,3,5,10,15,20天(长白落叶松)取样。取样时,每株自上到下摘取每个枝条上的完整针叶,每种处理3次重复,每5株松苗为1次重复,每次重复摘取鲜针叶10 g左右。每次从不同的样苗上采样,所采针叶充分混匀后放入冰箱冷冻保存,用于针叶内单宁含量的测定。
1.4 单宁含量测定 1.4.1 仪器LL-3000冷冻干燥机(英国基因有限公司)、电热恒温水浴锅(上海森信实验仪器有限公司)、奥豪斯电子天平(奥豪斯国际贸易上海有限公司)、752N紫外可见分光光度计(上海精密科学仪器有限公司)、D-37520冷冻离心机(德国)、冰箱(海尔有限公司)、烘箱(中外合资重庆四达实验仪器有限公司)。
1.4.2 试剂浓盐酸和甲醇(分析纯,天津市光复科技发展有限公司)、香草醛(分析纯,天津市光复精细化工研究所)、儿茶素、茉莉酸、茉莉酸甲酯和茉莉酮(色谱纯,Sigma公司)。
1.4.3 单宁含量测定参照徐伟(2006)的方法。求得标准曲线回归方程:Y=2.28X-0.004,R2=0.972 2。
1.5 数据处理与分析测得的数据应用Excel软件统计、处理,利用SPSS13.0软件以时间与茉莉酮(茉莉酸甲酯或茉莉酸)浓度2个因素,以及时间、外源物及外源物浓度3个因素,分别对兴安落叶松和长白落叶松处理数据进行单变量多因素方差分析(univariate analysis of variance)和Tukey HSD多重比较,对2种落叶松处理数据进行独立样本的t-test。
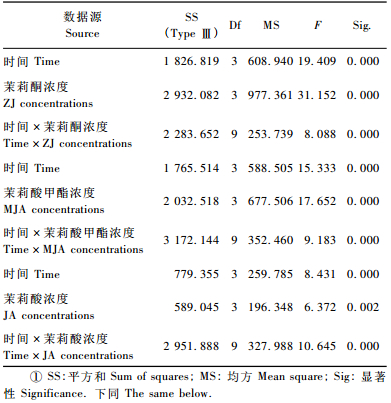
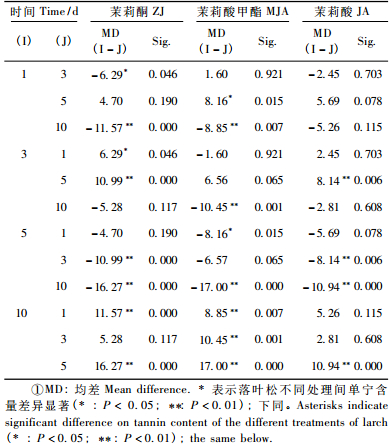
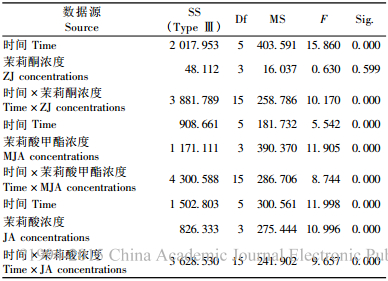
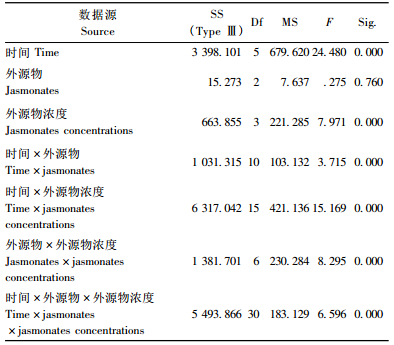
2 结果与分析 2.1 兴安落叶松针叶内单宁含量2因素方差分析由表 2看出,时间、茉莉酮(茉莉酸甲酯和茉莉酸)浓度对兴安落叶松针叶内单宁含量影响显著(P < 0.01)。图 1中4条不同浓度茉莉酮(茉莉酸甲酯或茉莉酸)曲线随处理时间变化相互交织在一起,表明时间与茉莉酮(茉莉酸甲酯或茉莉酸)浓度2因素的交互效应对兴安落叶松针叶内单宁含量影响显著(P < 0.01)(表 2)。处理后不同时间单宁含量变化的多重比较表明(表 3):茉莉酮处理1天与3, 10天,3天与5天,5天与10天; 茉莉酸甲酯处理1天与5, 10天,3天与10天,5天与10天; 茉莉酸处理3天与5天,5天与10天单宁含量差异显著(P < 0.05)。不同浓度外源物处理后多重比较发现(表 4),茉莉酮处理CK与0.01, 0.1, 1 mmol·L-1,0.01 mmol·L-1与0.1, 1 mmol·L-1; 茉莉酸甲酯处理CK与0.01 mmol·L-1,0.01 mmol·L-1与0.1, 1 mmol·L-1; 茉莉酸处理CK与1 mmol·L-1,0.01 mmol·L-1与0.1 mmol·L-1,0.1 mmol·L-1与1 mmol·L-1单宁含量有明显差异(P < 0.05)。
|
|

|
图 1 茉莉酮(a)、茉莉酸甲酯(b)、茉莉酸(c)浓度与时间交互效应对兴安落叶松针叶内单宁含量的影响 Figure 1 Effects of interactional effects of ZJ(a), MJA(b), JA(c) concentrations and time on tanin content in L.gmelinii needles |
|
|
|
|

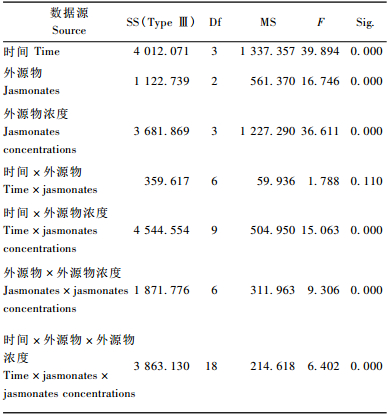
由表 5看出,时间、外源物、外源物浓度3因素对兴安落叶松针叶内单宁含量影响均显著(P < 0.01)。3因素对单宁含量影响的多重比较表明(表 6),处理后1天和5, 10天,3天和5, 10天,5天和10天单宁含量差异显著(P < 0.01);外源茉莉酮对兴安落叶松针叶内单宁含量的影响明显强于茉莉酸甲酯、茉莉酸(P < 0.01);而0.01, 0.1, 1 mmol·L-1浓度外源物对单宁影响明显强于CK,0.01 mmol·L-1浓度外源物对单宁影响显著大于其他浓度。
|
|
|
|
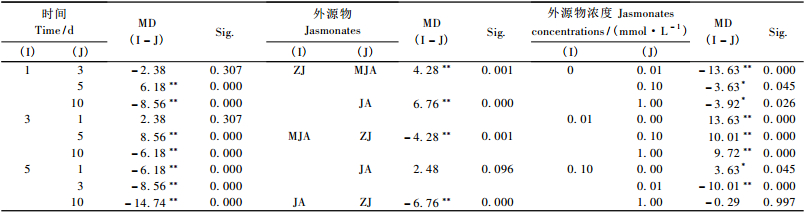
图 2a中3条外源物折线近似平行,说明时间与外源物间的交互作用对兴安落叶松单宁含量的影响不明显(P>0.05);图 2b中4条外源物浓度曲线随时间变化相互交叉,表明时间与外源物浓度对单宁含量的交互作用显著(P < 0.01);与图 2b相似,图 2c中3条外源物曲线随外源物浓度变化相互交叉,表示外源物与外源物浓度的交互作用明显影响兴安落叶松单宁含量(P < 0.01)(表 5)。图 1中4条不同浓度外源物曲线随处理时间变化相互交织在一起,表明时间、外源物与外源物浓度3个因素的交互效应对兴安落叶松针叶内单宁含量的影响极显著(P < 0.01)(表 5)

|
图 2 外源物与时间(a)、外源物浓度与时间(b)、外源物与外源物浓度(c)交线效应对兴安落叶松针叶内单宁含量的影响 Figure 2 Effects of interactional effects of jasmonates and time (a), jasmonats concentration and time (b) and jasmonates and jasmonates concentrations (c) on tannin content in L.gmelinii needles |
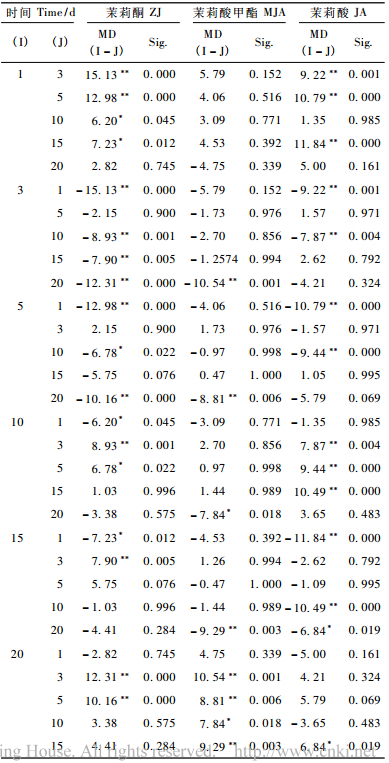
表 7中时间、茉莉酸甲酯(茉莉酸)浓度对长白落叶松针叶内单宁含量影响显著(P < 0.01)。图 3中4条不同浓度茉莉酮(茉莉酸甲酯或茉莉酸)曲线随处理时间的变化相互交织在一起,表明时间与茉莉酮(茉莉酸甲酯或茉莉酸)浓度2因素的交互效应对长白落叶松针叶内单宁含量有明显影响(P < 0.01)(表 7)。不同时间单宁含量的多重比较表明(表 8),茉莉酮处理1天与3, 5, 10, 15天,3天与10, 15, 20天,5天与10, 20天; 茉莉酸甲酯处理3天与20天,5天与20天,10天与20天,15天与20天; 茉莉酸处理1天与3, 5, 15天,3天与10天,5天与10天,10天与15天,15天与20天单宁含量差异显著(P < 0.05)。而不同外源物浓度的多重比较发现(表 9),不同浓度茉莉酮处理对单宁含量影响不显著(P>0.05);茉莉酸甲酯处理CK与1 mmol·L-1,0.01与0.1, 1 mmol·L-1; 茉莉酸处理CK与0.1 mmol·L-1,0.01与1 mmol·L-1,0.1与1 mmol·L-1单宁含量有明显差异(P < 0.05)。
|
|

|
图 3 茉莉酮(a)、苯莉酸甲酯(b)与茉莉酸(c)浓度与时间交互效应对长白落叶松针叶内单宁含量的影响 Figure 3 Effects of interactional effects of JA(a), MJA(b) and JA(c) concentrations and time on tannin content in L.olgensis needles |
|
|
|
|
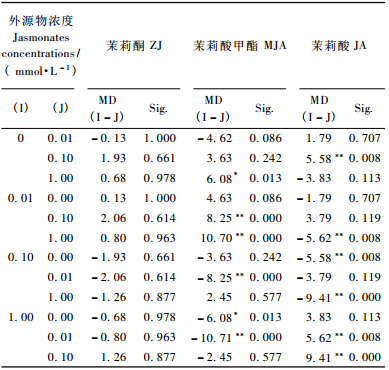
时间、外源物浓度对长白落叶松针叶内单宁含量影响极显著(P < 0.01)(表 10)。时间、外源物、外源物浓度对单宁含量影响的多重比较表明,处理后1天与3, 5, 15天,3天与10, 20天,5天与10, 20天,10天与15天,15天与20天单宁含量差异极显著(P < 0.01)(表 11); 不同外源物间对长白落叶松针叶内单宁含量的影响不显著(P>0.05)(表 12); 而外源物浓度处理CK与0.1 mmol·L-1,0.01与0.1 mmol·L-1,0.1与1 mmol·L-1对单宁影响有显著差异(P < 0.05)(表 12)。
|
|
|
|

|
|
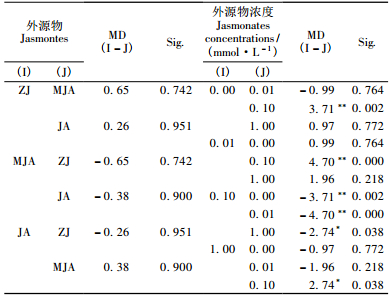
图 4中3条外源物曲线和4条外源物浓度曲线,随不同时间或不同外源物浓度波动并交织在一起,说明时间与外源物或外源物浓度,以及外源物和外源物浓度间的交互作用对长白落叶松单宁含量的影响明显(P < 0.01);图 3中4条不同浓度外源物曲线交织在一起,表明时间、外源物与外源物浓度3个因素对长白落叶松单宁含量的交互效应显著(P < 0.01)(表 10)。

|
图 4 外源物与时间(a)、外源物浓度与时间(b)、外源物与外源物浓度(c)交互效应对长白落叶松针叶内单宁含量的影响 Figure 4 Effects of interactional effects of jasmonates and time(a), jasmonates concentration and time(b), jasmonates and jasmonates concentrations(c) on tannin content in L.olgensis needles |
不同浓度外源茉莉酸处理后2种落叶松针叶内单宁含量相比,差异显著(表 13)。诱导后第1天,兴安落叶松处理ZJ0.1, MJA0.01, JA1针叶内单宁含量明显高于长白落叶松(PZJ0.1= 0.018,PMJA0.01=0.008,PJA1=0.003);而长白落叶松处理ZJ1、JA0.01针叶内单宁含量显著高于兴安落叶松(PZJ1=0.008,PJA0.01= 0.027)。第3天,兴安落叶松ZJ0.01, ZJ0.1, MJA0.1, JA0.01处理针叶内单宁含量明显高于长白落叶松(PZJ0.01=0.000,PZJ0.1=0.000,PMJA0.1= 0.047,PJA0.01=0.028)。第5天,兴安落叶松处理ZJ0.01针叶内单宁含量高于长白落叶松(PZJ0.01= 0.006);而长白落叶松处理MJA0.01针叶内单宁含量显著高于兴安落叶松(PMJA0.01=0.038)。第10天,兴安落叶松处理ZJ0.01, ZJ1, MJA0.01, MJA0.1, JA0.01针叶内单宁含量明显高于长白落叶松(PZJ0.01=0.014,PZJ1= 0.014,PMJA0.01=0.000,PMJA0.1=0.000,PJA0.01=0.021)。2种落叶松其他处理组针叶内单宁含量差异不显著(P>0.05)。
|
|
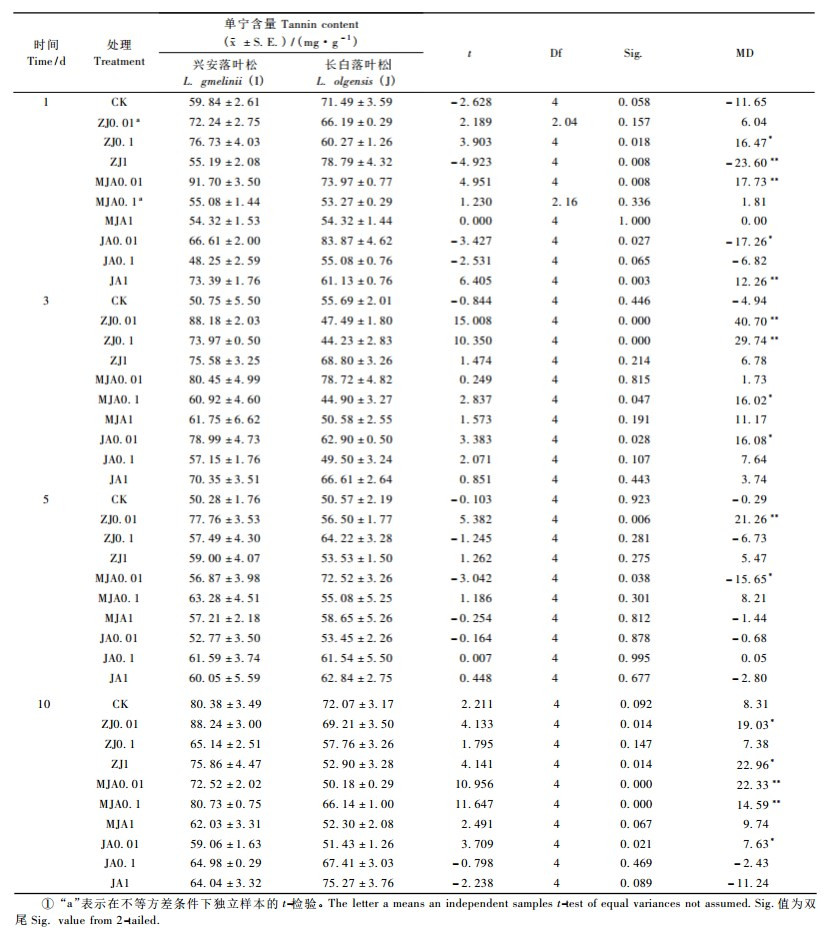
茉莉酸类化合物是一类重要的传递植物伤害信息的信号分子,是诱导植物产生直接或间接防御作用的关键性调节成分(Thaler et al., 2002; Cheong et al., 2003)。在施用外源茉莉酸类化合物后,2种落叶松针叶内单宁含量显著增加或减少,呈波动变化,诱导作用可延迟10~20 d。说明与机械伤害或昆虫取食一样(徐伟,2006; 袁红娥等,2009),外源茉莉酸类化合物能够诱导兴安和长白落叶松针叶内单宁含量显著变化,并具有诱导滞后性。但马尾松针叶损伤后,与对照相比,单宁含量在前期连续升高,而后期逐渐降低(王燕等,2001); 5年生兴安落叶松针叶被机械损伤或2年生兴安落叶松针叶受外源信号物质处理后,单宁含量一直高于对照(徐伟,2006; 袁红娥等,2009),此结果与本研究中单宁含量的变化差异较大,说明不同种植物或不同树龄及不同方法处理的同种植物对相同诱导的应激反应不同。
受害植物合成和积累单宁时,在代谢上是要花很大代价的。植物用于防御害虫的物质和能量是有限的,而植物本身又处于一种基础代谢与次生代谢交替开放的变化过程中,无论营养物质还是次生物质都处于不断的波动中(管致和,1996)。因此,不同处理时间的兴安和长白落叶松针叶内单宁含量呈现升高、降低的波动性变化,说明落叶松对生长和防御的兼顾性,其分配于两者的资源也处于动态变化过程中。
茉莉酮对兴安落叶松针叶内单宁含量的影响较另外2种外源物显著,3种外源物诱导长白落叶松针叶内单宁含量变化明显; 其中0.01 mmol·L-1茉莉酮、茉莉酸甲酯及1 mmol·L-1茉莉酸诱导兴安落叶松针叶内单宁含量增加显著,而0.01 mmol·L-1茉莉酸甲酯和1 mmol·L-1茉莉酸对长白落叶松针叶内单宁含量变化影响明显。表明落叶松对施用的外源茉莉酸类化合物具有明显的应激反应,且2种落叶松对不同种类或不同浓度的外源信号化合物不同。在诱导后第1, 3, 5和10天,兴安落叶松针叶内单宁含量显著高于长白落叶松(表 13),说明兴安落叶松对外源茉莉酸类化合物诱导作用的敏感性更强, 进一步证明了2种落叶松间防御反应的特异性。其他研究亦发现美洲栗(Castancea dentata)和板栗(C. mollissima)种间及组织间的组成单宁和诱导单宁存在明显的差异, 如果将2种栗子杂交, 将会改变其后代体内单宁的组成及含量, 影响其对病原物或植食动物的抗性(Kellogg et al., 2005; Cooper et al., 2008); 进一步用茉莉酸处理美洲栗, 增加其叶片中单宁的含量, 舞毒蛾(Lymantria dispar)幼虫取食该叶片后, 生长率较对照明显降低(Cooper et al., 2008)。王立春等(2008)用0.01, 0.1, 1 mmol·L-1茉莉酸甲酯喷雾处理的马尾松松针饲喂马尾松毛虫, 其中幼虫的取食量、排粪量、对食物的消耗和利用指数随着浓度的升高而降低, 而死亡率则相反; 任琴(2006)、邓文红等(2008)用茉莉酸甲酯熏蒸(或喷雾)和马尾松毛虫幼虫取食处理马尾松, 使其针叶内γ-氨基丁酸(G ABA)含量明显升高, 而游离氨基酸总量降低, 2种处理诱导马尾松释放萜类等挥发物的量和速度相似。这些研究均显示茉莉酸类化合物作为植物体内报警信号分子, 能够激活植物的防御系统, 诱导植物产生抗虫性。
在诱导防御中,单宁与其它代谢物质的关系如何?以及外源茉莉酸类化合物对中、壮龄落叶松体内单宁是否有同样的诱导作用?还有待于进一步研究。
程红, 严善春, 隋祥, 等. 2006. 黑龙江省主栽杨树品系干部单宁含量与青杨脊虎天牛危害的关系[J]. 东北林业大学学报, 34(2): 32-34. |
邓文红, 沈应柏, 李镇宇. 2008. 马尾松毛虫取食、茉莉酸甲酯和萜烯混合物处理对马尾松18种氨基酸含量的影响[J]. 林业科学, 44(10): 69-75. DOI:10.3321/j.issn:1001-7488.2008.10.012 |
戈峰, 李典漠, 邱业先, 等. 1997. 松树受害后一些化学物质含量的变化及其对马尾松毛虫种群参数的影响[J]. 昆虫学报, 40(4): 337-343. |
管致和. 1996. 植物医学导论[M]. 北京: 中国农业大学出版社: 193-205.
|
桂连友, 陈宗懋, 刘树生. 2005. 外源茉莉酸甲酯处理茶树对茶尺蠖幼虫生长的影响[J]. 中国农业科学, 38(2): 302-307. |
李水清, 张钟宁. 2007. 松墨天牛取食和人为损伤对马尾松针叶部分化学物质含量的影响[J]. 昆虫学报, 50(2): 95-100. |
李镇宇, 陈华盛, 袁小环, 等. 1998. 油松对赤松毛虫的诱导化学防御[J]. 林业科学, 34(2): 43-50. |
李镇宇, 王燕, 陈华盛, 等. 1999. 赤松毛虫的危害对小油松针叶内物质含量的影响[J]. 北京林业大学学报, 21(5): 41-45. |
任琴. 2006. 马尾松快速诱导抗性及化学信号物质的研究. 北京林业大学博士论文, 55-59, 86-96.
|
王燕, 戈峰, 李镇宇. 2001. 马尾松诱导化学物质变化的时空动态[J]. 生态学报, 21(8): 1256-1261. |
王立春, 任琴, 许志春, 等. 2008. 茉莉酸甲酯对马尾松松针萜烯类挥发物及马尾松毛虫生长发育的影响[J]. 北京林业大学学报, 30(1): 79-84. |
武予清, 郭予元. 2001. 棉花单宁-黄酮类化合物对棉铃虫的抗性潜力[J]. 生态学报, 21(2): 286-289. |
徐伟. 2006. 兴安落叶松诱导抗虫性研究. 东北林业大学博士论文, 39-42, 60-67.
|
薛皎亮, 谢映平, 刘计权, 等. 2000. 鞘蛾危害后诱导华北落叶松体内化学物质变化的研究[J]. 林业科学, 36(4): 46-50. DOI:10.11707/j.1001-7488.20000409 |
袁红娥, 严善春, 佟丽丽, 等. 2009. 剪叶及昆虫取食对兴安落叶松针叶中单宁的影响[J]. 生态学报, 29(3): 1415-1420. |
张时妙, 莫建初, 程梦林, 等. 2005. 单宁酸对淡色库蚊抗氰戊菊酯品系和敏感品系幼虫生长发育的影响[J]. 昆虫学报, 48(6): 886-891. |
Cheong J J, Choi Y D. 2003. Methyl jasmonate as a vital substance in plants[J]. Trends in Genetics, 19(7): 409-413. DOI:10.1016/S0168-9525(03)00138-0 |
Cooper W R, Rieske L K. 2008. Differential responses in American (Castanea dentata Marshall) and Chinese (C. mollissima Blume) chestnut (Falales:Fagaceae) to foliar application of jasmonic acid[J]. Chemoecology, 18: 121-127. DOI:10.1007/s00049-008-0399-y |
Gebauer R L E, Strain B R, Reynolds J F. 1998. The effect of elevated CO2 and N availability on tissue concentrations and whole plant pools of carbon-based secondary compounds in loblolly pine (Pinus taeda)[J]. Oecologia, 113: 29-36. |
Heijari J, Nerg A M, Kainulainen P, et al. 2005. Application of methyl jasmonate reduces growth but increases chemical defence and resistance against Hylobius abietis in Scots pine seedlings[J]. Entomologia Experimentalis et Applicata, 115: 117-124. DOI:10.1111/eea.2005.115.issue-1 |
Heyworth C J, Iason G R, Temperton V, et al. 1998. The effect of elevated CO2 concentrations and nutrient supply on carbon-based plant secondary metabolites in Pinus sylvestris L[J]. Oecologia, 115: 344-350. DOI:10.1007/s004420050526 |
Kellogg S K, Hebard F V, Rieske L K. 2005. Interactions between a fungal pathogen, foliar properties, and generalist herbivores[J]. Entomol Exp Appl, 117: 209-219. DOI:10.1111/eea.2005.117.issue-3 |
Muir A D, Gruber M Y, Hinks C F, et al. 1999. Effect of Condensed Tannins in the diets of major crop insects//Gross G G, Hemingway R W, Lew is N G. Plant Polyphenols-Chemistry, Biology, Phamacology, Ecology. New york:Kluwer Academic Publishers, 867-881.
|
Peters D J, Constabel C P. 2002. Molecular analysis of herbivore-induced condensed tannin synthesis:cloning and expression of dihydroflavonol reductase from trembling aspen (Populus tremuloides)[J]. Plant Journal, 32: 701-712. DOI:10.1046/j.1365-313X.2002.01458.x |
Pospisil J, Broche R G. 1981. Efectos del tanino sobre las larvas del mosquito Culex quinquef asciatus Say (Diptera:Culicidae). Informe Cientifico-Tecnico No.159. Instituto de Zoologia de la Academia de Ciencias de Cuba, La Habana. 8.
|
Räisänen T, Ryyppö A, Julkunen-Tiitto R, et al. 2008. Effects of elevated CO2 and temperature on secondary compounds in the needles of Scots pine (Pinus sylvestris L.)[J]. Trees, 22: 121-135. DOI:10.1007/s00468-007-0175-6 |
Scalbert A. 1991. Antimicrobial properties of tannins[J]. Phytochemistry, 30(4): 3875-3883. |
Seigler D S. 1998. Plant secondary metabolism[M]. Boston: Kluwer Academic Publishers: 6-9.
|
Thaler J S, Farag M A, Paré P W, et al. 2002. Jasmonate-deficient plants have reduced direct and indirect defences against herbivores[J]. Ecol Lett, 5: 764-774. DOI:10.1046/j.1461-0248.2002.00388.x |