 2010, Vol. 46
2010, Vol. 46文章信息
- 段爱国, 张建国, 何彩云, 曾艳飞
- Duan Aiguo, Zhang Jianguo, He Caiyun, Zeng Yanfei
- 干热河谷主要植被恢复树种干季光合光响应生理参数
- Characteristic Parameters of Light Response of Photosynthesis of Main Tree Species for Vegetation Restoration in Dry Season in Dry-Hot River Valley
- 林业科学, 2010, 46(3): 68-73.
- Scientia Silvae Sinicae, 2010, 46(3): 68-73.
-
文章历史
- 收稿日期:2009-02-11
-
作者相关文章


光是光合作用的主导因子,对每种绿色植物均可作出不同环境条件、不同发育阶段时的光合作用对光的响应曲线(沈允刚等,1998;张建国,2000)。光响应曲线模型是光合作用对环境因子响应模拟模型系统的核心组分,光响应曲线特征参数的确定,对了解光反应过程的效率非常重要(Robert et al., 1984)。通过光响应曲线可计算弱光条件下植物叶片的光合作用表观量子效率,有助于确定光合机构是否正常运转(陈根云等,2006), 而饱和光合强度可了解植物一定环境条件下的潜在光合作用能力。光响应曲线的拟合一直是国内外学者关注的焦点,一些机理模型与经验模型被研制并用于光合光响应曲线特征参数的求解,而众多研究实例表明这些光响应模型均具有各自不同不足之处。如通过直角或非直角双曲线模型途径所给出的光饱和点远小于实测值(刘宇峰等,2005;步秀芹等,2007),如何科学准确地求算光合光响应曲线特征参数是值得不断深入探究的问题。
干热河谷生态恶化,植被覆盖率低,水土流失严重,已经成为我国典型的生态脆弱区和造林困难地带,是全国生态环境建设的重点。干热河谷区气候干旱炎热,降雨量较少且分配不均,全年蒸发量为降水量的6倍左右,土壤干旱、贫瘠,耐干热、耐贫瘠树种的选择与造林是该区生态植被恢复的主要手段和途径。围绕干热河谷区生态植被的恢复,我国学者在立地分类评价(杨再强,2003)、造林树种选择(李昆等,2004)、造林技术(张信宝等,2001;林文杰等,2004)及水分生理(高洁,1997;马焕成等,2000;段爱国等,2007;2008)等方面开展大量的科学研究工作。但以往的研究对干热河谷特定生境条件下恢复造林树种的光合光响应特征参数缺乏足够探讨。本文以金沙江干热河谷13种植被恢复树种为试材,提出1种采用直线与抛物线相结合的分段函数拟合实测值并求算光合作用参数的方法,并创新性地提出1种科学界定低光强范围的方法,以期为光合光响应曲线特征参数的准确求解奠定理论与实践基础,并为干热河谷生境下植物适应性研究提供参考。
1 材料与方法 1.1 试验地概况试验区位于云贵高原云南省境内金沙江中段的元谋县。元谋县地处云南省北部,位于101°52′—102°00′ E,25°40′—26°00′N之间,海拔约1 100 m,属典型的干热河谷气候特征,雨季约5个月(6—10月),降水集中,但连续降雨日数较少,昼夜温差不大,空气闷热;干季6—7个月(11月至翌年5月),主要为3,4,5月,年降水量613.9 mm,其中6—10月份降雨占全年降水量的92%,年蒸发量3 847.8 mm,年均空气相对湿度53%。试验区土壤以燥红土为主,干旱瘠薄,水肥条件极差。自然植被以草丛为主,杂以灌木,稀少乔木,称为半自然稀树草原或稀树灌草丛。
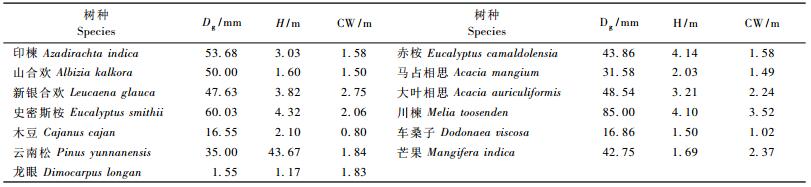
1.2 试验材料共选用13个野外试验树种,其中,印楝(Azadirachta indica)、新银合欢(Leucaena glauca)、木豆(Cajanus cajan)、马占相思(Acacia mangium)、大叶相思(Acacia auriculiformis)、史密斯桉(Eucalyptus smithii)、云南松(Pinus yunnanensis)、龙眼(Dimocarpus longan)及芒果(Mangifera indica)8个植被恢复造林树种均人工栽植于2002年,测定时树龄为5年,山合欢(Albizia kalkora)、车桑子及川楝等3个树种为野生树种,赤桉林为2002年砍伐后的萌生林分。所有供试树种均生长发育良好,各项生长指标如地径(Dg)、株高(H)、冠幅(CW)见表 1。
|
|
为避免因环境变化引起不同树种光合光响应曲线的不可比性,于2005年干季3月下旬,选择外界光照强度、温度及湿度均相对稳定的3个阴天,采用Licor-6400便携式光合测定系统测定13个树种的光合光响应曲线,重复1~2次。测定叶片均选取树冠中上部南向一级侧枝上的健康、成熟叶片。红蓝光源设置的光合有效辐射强度梯度为:1 800,1 600,1 200,800,500,200,100,50,20,0 μmol·m-2s-1,测定时间为9:25—15:50,大气CO2浓度约为(372±3) μmol·mol-1,温度为(29±1) ℃,空气相对湿度为28%±5%,测定前各树种均在1 200 μmol·m-2s-1光强下光诱导30 min。由于发现木豆在1 200 μmol·m-2s-1以上光强时即出现下降趋势,其光响应曲线最高光强设置为1 600 μmol·m-2s-1,并添加1 400 μmol·m-2s-1光强点。
1.3.2 光合作用光响应曲线拟合模型及相应光合生理参数求算作为1种经验模型,采用二次多项式即抛物线拟合的统计效果一般较好,并能计算出部分光合参数,如抛物线顶点的纵坐标及横坐标符合光响应曲线的最大净光合速率(Pmax)及光饱和点(LSP)的经验值,该方式无法得出表观量子效率,且会出现拟合所得的暗呼吸速率低于实测值,或为正值,而出现没有光补偿点的现象。为充分发掘抛物线方程精确、简明的优点,并合理求算出一般光合特征参数,笔者提出1种分段函数用以拟合光合光响应曲线,即将弱光阶段与强光阶段分别拟合,分段函数表达式如下所示:
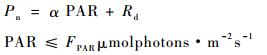
|
(1) |
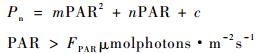
|
(2) |
上述分段函数中,FPAR为假定的弱光与强光分界点处的光强,参数α为植物表观光合量子效率,Rd为暗呼吸速率,并由(1)式可知光补偿点LCP为-Rd/α,由(2)式可得Pmax和LSP,这弥补了抛物线方程诸如无法得出表观量子效率、暗呼吸速率为正或没有光补偿点的缺陷,可准确反映弱光及强光阶段光合光响应曲线的特征,并得到相关的光合参数。相对应于弱光区域的表观光合量子效率,提出饱和光能利用效率(SLUE)的概念,用以描述饱和光强下单位光强的净光合速率,其定义式为:SLUE=Pmax/LSP。
1.3.3 弱光区域的确定方法通常弱光区域被界定为小于200 μmolphotons·m-2s-1,不同树种对光的需求程度是不同的,不同树种其弱光区域属性亦具有一定的差异。上述分段函数中,FPAR的确定最为关键,直接影响着表观量子效率的大小,亦关系着其他特征参数的求算。表观光合量子效率α的科学、准确确定有助于参数本身说理性的增强。此外,在估算LSP时直角双曲线及非直角双曲线方式均需借助弱光区域光合速率与光强的线性相关关系,弱光区域的合理确定实际上是不可回避且必须解决的问题。本文采用光合光响应曲线上光能利用效率下降前的光强范围作为植物弱光条件的具体界定。图 1为这种界定方法的示意图。

|
图 1 弱光区域界定方法的示意图 Figure 1 The sketch figure of method for determining the weak light region |
图 1中,(xi, yi)和(xi+1, yi+1)为线性方程上的2个点,α,β分别为点(xi, yi)和(xi+1, yi+1)与零点(0,0)相连后的直线与x轴的夹角。假设弱光条件下植物叶片表观光合速率随光照强度的增强而呈线性增大,则依图 2会存在tgβ>tgα,即yi+1/xi+1>yi/xi,这表明植物在(xi, yi)和(xi+1, yi+1)2点处的光能利用效率是递增的。反之,如植物在(xi+1, yi+1)处的光能利用效率低于在(xi, yi)处的光能利用效率,则表明植物在点(xi+1, yi+1)处时其表观光合速率已肯定不能维持线性增长,故以光合光响应曲线上光能利用效率的下降位点来界定植物表观光合速率随光强呈线性增长的弱光区域是可行的。这表明当在某处光强时光能利用效率出现第一次下降,则其前一处光强以内的光强范围即可当作植物的弱光区域。据此,图 1中FPAR的取值即为xi,弱光区域为[0, xi]。采用分段函数拟合方式,本文进一步比较分析了参数FPAR界定与否对供试树种光合光响应曲线特征参数求解值的影响。

|
图 2 13个树种光能利用效率随光强的变化趋势 Figure 2 The variation trend of light using efficiency of 13kinds of tree species with light intensity |
图 2描述了干热河谷13个树种光合光响应曲线上光能利用效率随光强的变化规律。从图 2可以看出:各树种光能利用效率随着光强的增强整体呈现先迅速升高而后逐渐下降的变化趋势,大致可分为快速上升, 下降, 平稳3个阶段。印楝、川楝、马占相思、山合欢、新银合欢、车桑子、赤桉、云南松及龙眼等9个树种的光能利用效率在光强为200 μmolphotons·m-2s-1时出现第一次下降,木豆、大叶相思及芒果等3个树种在光强为100 μmolphotons·m-2s-1时出现第一次下降,仅史密斯桉在光强达到200 μmolphotons·m-2s-1后其光能利用效率才出现下降。这表明史密斯桉在200 μmolphotons·m-2s-1后其表观光合速率始不能维持线性增长,而木豆、大叶相思及芒果在50 μmolphotons·m-2s-1后表观光合速率已不能维持线性增长,其他树种在100 μmolphotons·m-2s-1后表观光合速率不能维持线性增长。由此可知,史密斯桉的弱光区域为[0, 200 μmolphotons·m-2s-1],印楝、川楝、马占相思、山合欢、新银合欢、车桑子、赤桉、云南松、龙眼等树种的弱光区域为[0, 100 μmolphotons·m-2s-1],而木豆、大叶相思及芒果的弱光区域仅为[0, 50 μmolphotons·m-2s-1]。可以发现,绝大多数树种的弱光区域实际上均小于通常认为的200 μmolphotons·m-2s-1。供试树种光能利用效率随光强的这种变化规律为采用光能利用效率下降位点界定弱光区域提供了实践基础,而各树种弱光区域的界定为进一步准确求算光合光响应曲线特征参数提供了科学基础。
2.2 弱光区域的界定对光合光响应曲线特征参数的影响从表 2可以看出:分段函数对供试树种光合光响应曲线具有良好的拟合性能,拟合相关系数均在0.9以上。弱光区域界定与否对供试树种光合光响应曲线的特征参数产生了较为明显的影响。对13个供试树种同一特征参数在弱光区域界定前后的求解值进行双因素方差分析,结果表明:除去界定前后弱光区域表现一致的树种外,弱光区域的界定与否对表观量子效率α和暗呼吸速率Rd均具极显著影响(概率P < 0.01),而对光补偿点LCP、光饱和点LSP及最大表观光合速率Pmax等3个特征参数影响不显著,概率P值分别为0.439 2,0.324 0,0.358 9。分树种对弱光区域属性进行分别界定后,绝大部分供试树种的表观量子效率均有所提高,而暗呼吸速率有所降低,这表明当不分树种统一采用200 μmolphotons·m-2s-1作为分界定点时,求解得到的表观量子效率及暗呼吸速率一般分别表现为低估与高估。弱光区域的界定与否对不同树种光合特征参数估计值的影响程度是不同的。
|
|
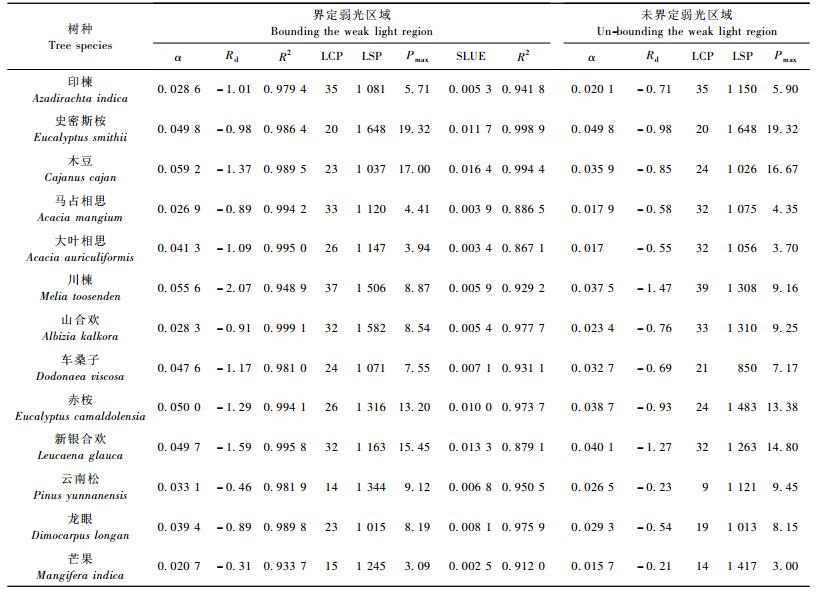
鉴于以光合光响应曲线上光能利用效率的下降位点来界定植物表观光合速率随光强呈线性增长的弱光区域更符合不同树种对低光需求的差异性特性,以对弱光进行界定后采用分段函数拟合所得的光响应曲线特征参数为对象,探讨干热河谷植被恢复树种在干季3月的光合生理特性。
2.3.1 表观光合量子效率表观光合量子效率(α),又称低光强下的光合量子效率,是反映植物光能利用和光合物质生产效率的基本参数,是构建冠层光合作用模拟模型及C循环模型的重要生理指标。从表 2可知:元谋干热河谷13个树种在干季3月的表观光合量子效率大小范围为0.020 7~0.059 2 μmol CO2·m-2s-1/μmolphotons·m-2s-1。各树种表观光合量子效率大小比较结果为:木豆>川楝>赤桉>史密斯桉>新银合欢>车桑子>大叶相思>龙眼>云南松>印楝>山合欢>马占相思>芒果,表观光合量子效率越大说明树种在低光强阶段的光能利用效率愈高。
2.3.2 暗呼吸速率与光补偿点由表 2可知:干热河谷植被恢复树种具有不同程度的暗呼吸速率,总体范围为-2.07~ -0.31 μmol CO2·m-2s-1。可以发现,供试树种的暗呼吸速率指标与表观光合量子效率指标存在一定的正相关关系,即树种表观光合量子效率越大,其暗呼吸速率越强。供试树种光补偿点范围在14~37 μmolphotons·m-2s-1,均小于50 μmolphotons·m-2s-1,故在测试光合光响应曲线时,为准确得到树种光补偿点,有必要在0~50 μmolphotons·m-2s-1之间设置1~2个测点。
2.3.3 光饱和点由表 2可知:13个供试树种在干季3月的光饱和点的大小范围为1 015~1 648 μmolphotons·m-2s-1,由高到低依次为史密斯桉、山合欢、川楝、云南松、赤桉、芒果、新银合欢、大叶相思、马占相思、印楝、车桑子、木豆及龙眼。按喜光程度划分,可大致分为3类:史密斯桉、山合欢、川楝的光饱和点最高,属强度喜光树种;云南松、赤桉和芒果居中,属中度喜光树种;新银合欢、大叶相思及木豆等7个树种属弱度喜光树种。元谋干热河谷除温度高、空气湿度低、土壤贫瘠外,干热季节典型晴天里光照强度较高亦是1个显著的特征,其典型晴天里的最大光照强度一般均在1 600 μmol·m-2s-1以上,几乎高于所有供试树种的光饱和点,故就光照强度1个因子而论,干季典型晴天里各树种均有可能发生光抑制现象。
各树种饱和光能利用效率SLUE大小排序为:木豆>新银合欢>史密斯桉>赤桉>龙眼>车桑子>云南松>川楝>山合欢>印楝>马占相思>大叶相思>芒果,饱和光能利用效率越高,树种利用光能合成有机物质的能力则越强。可以看出:各树种饱和光能利用效率大小排序与表观光合量子效率大小排序具有一定的相似性,这表明在弱光区域光能利用效率高的树种,其在达到光饱和点时的光能利用效率一般亦越高。
2.3.4 最大净光合速率由表 2可知:干热河谷植被恢复树种的最大净光合速率呈现出明显的差异,取值在3.09~19.32 μmol CO2·m-2s-1间变动,变化幅度较大。13个树种最大净光合速率由大至小依次为:史密斯桉、木豆、新银合欢、赤桉、云南松、川楝、山合欢、龙眼、车桑子、印楝、马占相思、大叶相思、芒果。Pmax越大,表明树种在干热河谷这一特定生境条件下的潜在光合能力越强。此外,可以发现Pmax,α,SLUE 3者间的树种排序比较相似,这表明树种在弱光区域及饱和光强下的光能利用效率愈高,其最大净光合效率一般亦越高。
3 结论与讨论金沙江干热河谷元谋段13种野外生长树种在干季的光能利用效率随光强的变化规律表明:以光合光响应曲线上光能利用效率的下降位点来界定植物表观光合速率随光强呈线性增长的弱光区域具有理论与实践基础,是科学可行的,避免了弱光区域选取的人为性,且各树种弱光区域的具体界定为进一步准确求算光合光响应曲线特征参数提供了科学基础。为精确得到光响应曲线光能利用效率的下降位点,进行植物光合光响应曲线测定时,在0~200 μmolphotons·m-2s-1间按一定的递增模式多设置光强点是十分必要的。鉴于在估算LSP时直角双曲线及非直角双曲线方式均需借助弱光区域光合速率与光强的线性相关关系(刘宇峰等,2005;步秀芹等,2007),弱光区域的具体界定方法可在光响应曲线的拟合及特征参数的求算领域得到广泛的应用,同时将有助于不同植物种类、不同试验条件下植物表观光合量子效率的合理求算,并对光合光响应曲线发展阶段的量的划分及理解具有一定的理论与实践意义。
1条完整的植物叶片光合作用光响应曲线一般可以划分为3个不同的发展阶段:1)弱光条件下净光合速率随光强线性增加的阶段;2)净光合速率以递减的增速随光强上升的阶段;3)始于光饱和点的不同程度的光抑制阶段。应该说整个过程并不复杂,对其模拟的难点主要在于模型参数的说理性及准确性的同时获得,而目前广泛应用的各种模型均很难兼顾。基于光响应曲线特征参数的重要性及准确求算的必要性,本文采用分段模拟的思路,充分利用抛物线方程的优点(郭志达等,1999),结合弱光阶段的具体界定给出了1种较好的解决途径,试验结果表明分段函数对供试树种光合光响应曲线具有良好的拟合性能。
弱光区域的界定与否对表观光合量子效率α和暗呼吸速率Rd均具极显著影响,而对光补偿点LCP,光饱和点LSP及最大表观光合速率Pmax等3个特征参数影响不显著,这亦反映光响应曲线拟合中弱光区域界定的作用及必要性。
元谋干热河谷13个树种在干季3月的表观光合量子效率、暗呼吸速率、光补偿点、光饱和点、最大净光合速率大小范围分别为0.020 7~0.059 2 μmol CO2·m-2s-1/μmolphotons·m-2s-1,-2.07~-0.31 μmol CO2·m-2s-1,14~37 μmolphotons·m-2s-1,1 015~1 648 μmolphotons·m-2s-1,3.09~19.32 μmol CO2·m-2s-1,其中表观光合量子效率的大小均不同程度低于自然界植物最大量子效率的理论值0.08~0.125 μmol CO2·m-2s-1/μmolphotons·m-2s-1(Long et al., 1994)。由于受到多树种平行比较的限制,每一树种光合光响应曲线测定的重复数略显不足,故涉及树种特征参数大小比较的排序结果仅供参考,尚待今后针对部分树种的试验予以验证或修正。而对于干热河谷植被恢复树种系统性的光合光响应特性研究,亦尚需进一步展开各树种光合光响应曲线的土壤水分、大气湿度、温度及综合效应的探讨,这种深入的研究有望从光合生理的角度对树种的生态适应性给出满意的解释。
步秀芹, 徐学选, 康博文, 等. 2007. 黄土丘陵区铁杆蒿光合特性[J]. 林业科学, 43(9): 623-628. |
陈根云, 俞冠路, 陈悦. 2006. 光合作用对光和二氧化碳响应的观测方法[J]. 植物生理与分子生物学学报, 32(6): 691-696. |
段爱国, 张建国, 张俊佩, 等. 2007. 金沙江干热河谷主要植被恢复树种叶水势的时空变化规律[J]. 林业科学研究, 20(2): 151-159. |
段爱国, 张建国, 何彩云, 等. 2008. 干旱胁迫下金沙江干热河谷主要造林树种盆植苗的蒸腾耗水特性[J]. 林业科学研究, 21(4): 436-445. |
高洁. 1997. 元谋干热河谷主要造林植物的耐旱性评估[J]. 西南林学院学报, 17(2): 19-23. |
郭志达, 张宏达, 李志安, 等. 1999. 鹅掌楸苗期光合特性的研究[J]. 生态学报, 19(2): 164-169. |
马焕成, 胥辉, 陈德强. 2000. 元谋干热河谷几种相思和桉树水分消耗量估测[J]. 林业科技通讯, (4): 9-11. |
李昆, 张春华, 崔永忠, 等. 2004. 金沙江干热河谷区退耕还林适宜造林树种筛选研究[J]. 林业科学研究, 17(5): 555-563. |
林文杰, 马焕成, 周蛟. 2004. 干旱胁迫下不同保水剂处理的水分动态研究[J]. 水土保持研究, 11(2): 121-123. |
刘宇锋, 萧浪涛, 童建华, 等. 2005. 非直线双曲线模型在光合光响应曲线数据分析种的应用[J]. 中国农学通报, 121(8): 76-79. |
沈允钢, 施教耐, 许大全. 1998. 动态光合作用[M]. 北京: 科学出版社.
|
杨再强, 王立新, 潘步昌, 等. 2003. 攀西干旱干热河谷退耕还林立地类型的划分及经营模式探讨[J]. 四川林勘设计, 2(2): 33-35. |
张建国, 李吉跃, 沈国舫. 2000. 树木耐旱特性及其机理研究[M]. 北京: 中国林业出版社.
|
张信宝, 杨忠, 文安邦. 2001. 微水造林建设攀枝花市视野区常绿森林植被[J]. 水土保持学报, 15(4): 6-9. |
Long S P, Humphries S, Falkowski P G. 1994. Photo inhibition of photosynthesis in nature[J]. Annu Rev Plant Physiol Mol Biol, 45: 633-662. DOI:10.1146/annurev.pp.45.060194.003221 |
Robert E S, Mark A, John S B. 1984. Kok effect and the quantum yield of photosynthesis[J]. Plant Physiology, 75: 95-101. DOI:10.1104/pp.75.1.95 |