 2010, Vol. 46
2010, Vol. 46文章信息
- 同小娟, 张劲松, 孟平, 黄辉, 国琳, 尹昌君, 高峻
- Tong Xiaojuan, Zhang Jinsong, Meng Ping, Huang Hui, Guo Lin, Yin Changjun, Gao Jun
- 华北低丘山地人工混交林净生态系统碳交换的变化特征
- Characteristics of Net Carbon Exchange over a Mixed Plantation in a Hilly Area of the North China
- 林业科学, 2010, 46(3): 37-43.
- Scientia Silvae Sinicae, 2010, 46(3): 37-43.
-
文章历史
- 收稿日期:2009-07-16
-
作者相关文章



2. 中国林业科学研究院林业研究所 国家林业局林木培育重点实验室 北京 100091
2. Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration Research Institute of Forestry, CAF Beijing 100091
在全球碳循环研究中,人们不仅关注不同陆地生态系统的碳储量,而且更关注不同生态系统碳储量变化(碳源/碳汇)在时间和空间上的分布特征,以及这种特征对全球变化的响应和适应。认识和理解不同陆地生态系统碳源和碳汇功能时空变化的有效方法之一是直接测算陆地植被与大气间的碳通量。涡度相关法被认为是长期测算生态系统碳通量最可靠和切实可行的方法(Baldocchi et al., 1996; Oechel et al., 2000)。由于森林在陆地生态系统碳循环和全球变化中起着举足轻重的作用,许多国家将陆地碳通量观测和研究的重点放在森林方面。
我国于2002年开始采用涡度相关技术对森林水、碳通量进行观测,但主要集中于天然林生态系统(关德新等, 2004; 张一平等, 2006; 吴家兵等, 2006; 2007; Zhang et al., 2006; Guan et al., 2006)。近20年来,全国森林碳储量增加了约0.4 Pg C,年增加0.011~0.035 Pg C。森林碳储量的增加主要是人工造林的结果(Fang et al., 2001)。我国人工林面积已占森林总面积的30%, 人工林潜在的碳汇功能不可低估。以往对人工林碳平衡的研究大多采用生物量调查法,而基于涡度相关法的碳交换研究则较少。从已有文献看,我国对人工林碳通量的观测和研究主要在温带(Wang et al., 2008)和亚热带(刘允芬等, 2004;2006; Yu et al., 2008; 彭镇华等, 2009),有关其他气候带的研究还未见报道。人工林碳通量数据的缺乏势必影响我国森林碳收支评价的准确性。
华北山地作为华北平原重要的生态屏障,一直是我国林业生态建设的重点区域。对该地区人工林生态系统碳收支状况进行综合研究,可填补我国在暖温带人工林碳交换研究的空白,对评价林业生态工程对碳循环和气候变化的影响具有重要意义。
1 试验区概况和研究方法 1.1 试验区概况试验在黄河小浪底森林生态系统定位研究站进行(35°01′ N,112°28′ E,平均海拔410 m)。该站位于河南省济源市,面积7 210 hm2,地处黄河中游流域,紧连太行山,属暖温带亚湿润季风气候,4—9月盛行东北风。年平均气温12.4~14.3 ℃,全年日照时数为2 367.7 h,年平均降水量641.7 mm。受季风气候的影响,降水季节性分配不均匀。6—9月平均降水量为438.0 mm,占全年的68.3%。主要树种为栓皮栎(Quercus variabilis)、侧柏(Platycladus orientalis)、刺槐(Robinia pseudoacacia)等,林龄分别为32,30和28年,平均株高分别为10.5,8.2和9.3 m。土壤种类主要为棕壤和石灰岩风化母质淋溶性褐土。
通量观测塔位于站区1号样地内,海拔412 m。塔周围1.8 km2范围内平均坡度约14°,林木覆盖率96.0%左右。主要树种为栓皮栎、侧柏、刺槐,所占比例分别约为80%,8%和12%,呈团状混交。
1.2 试验观测涡度相关系统安装在观测塔的东北方向,高度为30.0 m。涡度相关系统包括CSAT3三维超声风速仪(Campbell Sci. Inc., USA)和LI-7500开路式红外CO2/H2O分析仪(Li-Cor Inc., USA)。原始数据由CR5000数据采集器(Campbell Sci. Inc., USA)采集,采样频率为10 Hz。
微气象梯度观测系统包括7层AR-100风速仪(Vector Instruments, UK)和7层HMP45C型温湿度传感器(Vaisala, Helsinki, Finland)(安装高度分别为8, 9, 11, 14, 18, 26 m)。在27 m高度处安装了CNR-1型净辐射表和CM11型总辐射表(Kipp and Zonen)。此外,还安装Li-190SB型点状光量子表(Li-Cor Inc., USA)、气压计和雨量计(Model 52203, RM Young, Inc.)等。上述仪器均与CR23x型数据采集器(Campbell Sci., USA)相连,每30 min输出1组平均值。
观测塔附近地表下2 cm处安装了土壤热流板(Model HFP01SC, Hukseflux, Netherland),土壤温度传感器(107 L, Campbell Sci. Inc.)分别安装于地表及地下5, 10, 20和40 cm。土壤水分由ECH2O型(Decagon Devices,USA)土壤水自动观测系统测定,观测深度为10,20和40 cm。
1.3 涡度相关数据处理对涡度相关测定的10 Hz原始数据进行处理,得到三维风速、温度、湿度、CO2浓度和气压的30 min平均值和协方差。CO2通量由垂直风速(w)与CO2浓度(c)脉动值的协方差w′c′计算获得。经3次坐标旋转(Wilczak et al., 2001)和WPL校正(Webb et al., 1980)后得到30 min平均CO2通量。
观测中夜间湍流较弱时,从土壤和植物冠层释放的CO2难以到达仪器测定高度,仪器观测精度降低会导致夜间CO2通量偏低。对夜间低于临界摩擦风速(U*)时所得数据应当剔除。另外,仪器故障、人为影响以及雨天或清晨有露水会导致错误数据的出现,对这些异常值应进行删除。对经过初步筛选的数据分白天和黑夜计算月平均值和方差,与平均值相差超过3倍方差的数值也被认为是异常点。对上述被去除的数据和由于仪器故障、停电等原因丢失的数据用下列方法进行插补:1)小于2 h的缺失数据用线性内插法插补;2)大于2 h的缺失数据采用平均日变化法(mean diurnal variation, MDV)进行插补(Falge et al., 2001)。
1.4 净生态系统碳交换的计算净生态系统碳交换(net ecosystem carbon exchange, NEE)由下式给出(Aubinet et al., 2000):
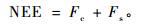
|
(1) |
式中: Fc为CO2湍流通量(mg·m-2s-1),由涡度相关法观测获得; Fs为涡度相关仪器观测高度以下大气CO2储存通量(mg·m-2s-1),可由下式进行估算(Hollinger et al., 1994; Greco et al., 1996):
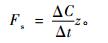
|
(2) |
式中: ΔC为高度z处前后2次相邻时间测定的CO2浓度差(mg·m-3),Δt为前后2次测定的时间间隔(s),z为CO2浓度观测高度(m)。NEE为负时表示人工林净吸收大气CO2,反之表示净排放CO2。
2 结果与讨论 2.1 夜间CO2通量与摩擦风速的关系在夜间大气层结稳定条件下,湍流交换较弱,涡度相关仪器测定的CO2通量往往偏低(Baldocchi, 2003)。在碳通量研究中,通常是根据摩擦风速(U*)与CO2通量的关系确定一个临界U*(Falge et al., 2001),摩擦风速小于临界U*时所得的CO2通量数据则被剔除掉。由人工林2006和2007年全年夜间半小时CO2通量和U*的关系可知,当U*小于0.2 m·s-1时,CO2通量随U*的增加而增大; 当U*超过0.2 m·s-1后,CO2通量趋向稳定(图 1)。因此,在本研究中,U*小于0.2 m·s-1时对应的CO2通量数据将被舍去。这与刘允芬等(2004)、关德新等(2004)、Wang等(2008)在中亚热带针叶林、温带阔叶红松(Pinus koraiensis)林和温带落叶林所得结果类似。

|
图 1 人工混交林夜间CO2通量与摩擦风速(U*)的关系 Figure 1 Relationship between nighttime CO2 fluxes with friction velocity (U*) in a mixed plantation |
研究表明,生长季(4—10月)人工混交林NEE具有明显的日变化规律(图 2)。日出后,随着辐射和气温的增加,光合作用逐渐加强,人工混交林由释放CO2转而吸收CO2,NEE由正变负。至12: 00前后,人工混交林的CO2吸收量达到最大值。14: 00以后,随着辐射和气温下降,光合作用逐渐变弱,人工混交林的CO2吸收量也逐渐减少。日落前后,人工混交林由吸收CO2转而释放CO2,NEE由负变正。夜间人工混交林的CO2释放量在小范围内波动,并明显少于白天CO2的吸收量。

|
图 2 人工混交林NEE月平均日变化 Figure 2 Diurnal variation of monthly mean net ecosystem carbon exchange in a mixed plantation |
在植物生长季,不同月份人工混交林的NEE日变化规律大体类似,但也有所不同。2006和2007年月平均日最大CO2吸收量分别以5月和7月最大(均为-0.86 g CO2·m-2d-1),而都以10月最小。月平均NEE日较差则以7, 8月最大。每日人工混交林吸收CO2的起始时间以5, 6月最早(约7: 00),10月最晚(约9: 00);每日人工混交林吸收CO2的结束时间4—7月都在19: 00左右,8月后逐渐提前,10月提前至5: 30。每日人工混交林吸收CO2的持续时间以5, 6月最大(12 h),10月最小(8.5 h)。
值得注意的是,当地6月的辐射和气温都是全年最高或接近最高的,但人工混交林的CO2吸收量无论日最大值还是日平均值都小于相邻其他月份。究其原因,主要是由于空气干燥,空气饱和差(vapour pressure deficit, VPD)较大,叶片的气孔阻力较大,从而抑制了植物的光合作用。NEE日较差主要受气温影响,一般气温高NEE的日较差较大; 但也有例外,如前所述,2006年6月气温虽高,但VPD很大,CO2的日最大吸收量较小,NEE日较差也较小。
进入非生长季(11月至翌年3月),随着气温下降,除少量针叶外,大部分树叶枯黄凋落。加之辐射和气温都很低,生态系统的光合作用与呼吸作用很小,人工混交林与大气间的CO2交换十分微弱,NEE日变化不明显。
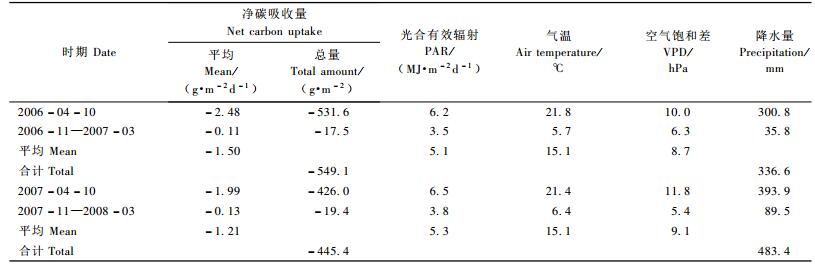
2.3 净生态系统碳交换季节变化受温度、辐射和降水等因素影响,人工混交林NEE的季节变化较大(图 3)。2006和2007年日平均NEE变化范围分别为-27.1~8.1和-24.4~9.8 g CO2·m-2d-1。2006年初春气温回升快,树叶萌发早,且春季降水满足了树木生长需要,人工混交林生长迅速。年最大日平均CO2吸收量出现于5月10日,平均CO2吸收量最大的月份也是5月。夏季受干旱影响,人工混交林的CO2吸收时有升降,但随时间推移呈总体下降趋势(图 3, 4)。2007年春季气温回升略迟于2006年, 虽然3月降水较多,但4—5月降水只有2006年同期的一半。干旱胁迫导致叶片气孔阻力增大,植物光合作用受到抑制, 树木的生长发育受到影响。春季人工混交林的CO2吸收量明显低于2006年同期(图 4)。入夏,较多的降雨使干旱得到缓解,树木生长迅速。年最大日平均CO2吸收量迟至8月20日出现, 之后人工混交林的CO2吸收量迅速下降,于9月中旬达到低谷(图 3)。由于8月上半月人工混交林CO2吸收低谷的存在,平均CO2吸收量最大的月份并非8月,而是此前的7月(图 4)。10月以后,随着气温下降,树叶枯黄凋落,人工混交林叶面积指数迅速降低, 冬季人工混交林只有少量针叶,叶面积很小, 加之温度、辐射很低,生态系统的光合作用与呼吸作用都很弱。人工混交林与大气间的碳交换在小范围内波动,净碳吸收量很小(图 3, 4)。分析表明,生长季人工混交林的碳汇强度为非生长季的15~22倍,生长季累计净碳吸收占全年的96%左右(表 1)。

|
图 3 人工林NEE季节变化(2006—2007年) Figure 3 Seasonal variation of net ecosystem carbon exchange in a mixed plantation from 2006 to 2007 |

|
图 4 人工林月平均NEE、光合有效辐射、气温、空气饱和差和降水量的季节变化 Figure 4 Seasonal variations of monthly mean net ecosystem carbon exchange, photosynthetically active radiation(PAR), air temperature(T), vapour pressure deficit (VPD) and precipitation (P) in a mixed plantation from 2006 to 2007 |
|
|
值得注意的是,2年的6月辐射和温度都是全年最高或接近最高的,但NEE却较低。除降水少、土壤干旱影响树木生长外,2年6月较高的VPD也是重要原因。土壤干旱及空气干燥使气孔阻力增大,抑制了光合作用,导致人工混交林的CO2吸收量变小。
2.4 年碳收支观测结果表明: 2006和2007年人工混交林的年净碳吸收量分别为549.1和445.4 g C·m-2a-1,平均为497 g C·m-2a-1(表 1)。人工混交林是大气较强的碳汇,且年际变化显著。与2006年相比,2007年辐射略高,降水高出44%,而净碳吸收量反比2006年低了近20%(表 1)。究其原因,主要是降水分配不均引发季节性干旱所致。2007年年降水量虽然高出2006年近一半,但春季干旱严重,4,5月降水量只有2006年同期的一半(图 4)。干旱胁迫导致叶片气孔阻力增加,光合作用受到抑制, 树木生长缓慢,叶面积较小。2006年4, 5月正是全年辐射最高的时候,由于干旱人工混交林比2006年同期少吸收了约130 g C·m-2的CO2。虽然2007年夏季雨水充沛,人工混交林的净碳吸收高于2006年同期(图 4),但也难以补回春季的亏空。春季干旱是2007年人工混交林年净碳吸收显著下降的主要原因。刘允芬等(2006)发现,夏季干旱胁迫使中亚热带人工林生态系统的碳吸收减少了9.5%。本试验中,春季干旱的影响更大,干旱胁迫使暖温带人工混交林生态系统的碳吸收下降了19%。
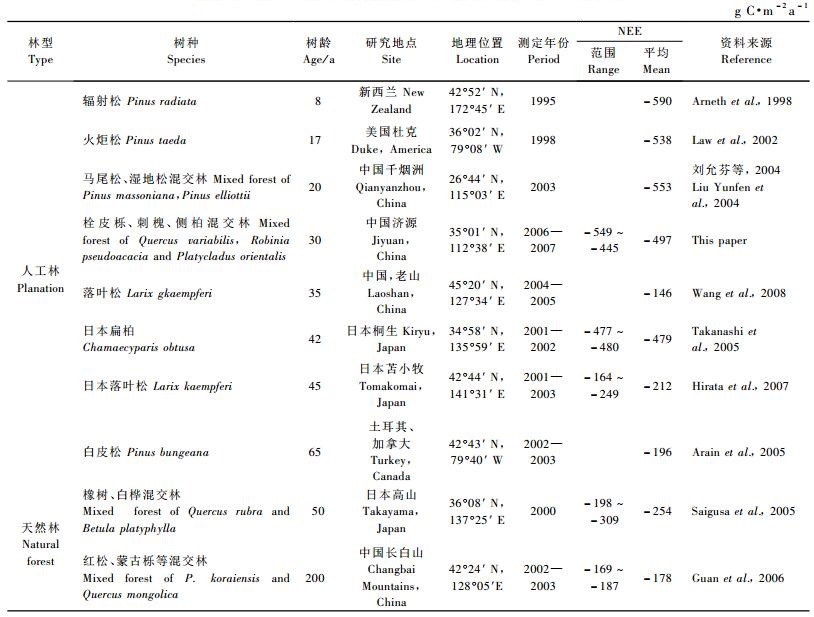
表 2列出了文献报道的从26°-45° N共10个站点的碳通量观测结果,涵盖从亚热带、暖温带到温带不同树种的人工林和天然林生态系统。本文测定结果在人工林的碳通量范围之内,而高于天然林的碳通量观测值(表 2)。从表 2可以看出,人工林的NEE变化范围很大,从-146到-590 g C·m-2a-1不等。不同地区气候、土壤及树种差别是导致森林碳汇功能差异的重要原因。与天然林相比,人工林的林龄一般较低,具有生长迅速、固碳能力强的特点。若不同森林生态系统在气候、土壤及树种甚至林型上的差别,仅分析NEE与林龄的关系,则可发现碳吸收与林龄呈显著的负相关关系(P < 0.01)。相对于天然林,林龄较低的人工林在固碳方面无疑有着巨大的优势。
|
|
通过在华北低丘山地30年生栓皮栎-刺槐-侧柏人工混交林连续2年的碳通量观测,得出以下结论:
1) 人工混交林NEE的日变化在生长季(4—10月)十分显著,在非生长季(11月至翌年3月)不明显。2006和2007年月平均日最大CO2吸收量分别以5, 7月最大。月平均NEE日较差则以7,8月最大。
2) 2006和2007年人工混交林的日平均NEE变化范围分别为-27.1~8.1和-24.4~9.8 g CO2·m-2d-1,季节变化明显。2006和2007年月平均CO2吸收量最大的月份分别为5, 7月。生长季累计净碳吸收占全年的96%左右。
3) 2006和2007年人工混交林的年净碳吸收量分别为549.1和445.4 g C·m-2a-1。人工混交林是大气较强的碳汇,且年际变化显著。春季干旱是2007年人工混交林净碳吸收显著下降的主要原因。鉴于华北低丘山地人工混交林生态系统碳交换年际波动较大,今后还需要进行长期的碳通量观测,深入探讨人工混交林净碳交换对各生物及环境因子的响应机制,为准确评价我国森林碳汇提供理论依据和数据支持。
关德新, 吴家兵, 于贵瑞, 等. 2004. 气象条件对长白山阔叶红松林CO2通量的影响[J]. 中国科学D辑:地球科学, 34(增刊Ⅱ): 103-108. |
刘允芬, 宋霞, 孙晓敏, 等. 2004. 千烟洲人工针叶林CO2通量季节变化及其环境因子的影响[J]. 中国科学D辑:地球科学, 34(增刊Ⅱ): 109-117. |
刘允芬, 于贵瑞, 温学发, 等. 2006. 千烟洲中亚热带人工林生态系统CO2通量的季节变异特征[J]. 中国科学D辑:地球科学, 36(增刊Ⅰ): 91-102. |
彭镇华, 王妍, 任海青, 等. 2009. 安庆杨树林生态系统碳通量及其影响因子研究[J]. 林业科学研究, 22(2): 237-242. |
吴家兵, 关德新, 孙晓敏, 等. 2006. 长白山阔叶红松林主要树种及群落冠层光合特征[J]. 中国科学D辑:地球科学, 36(增刊Ⅰ): 83-90. |
吴家兵, 关德新, 张弥, 等. 2007. 长白山阔叶红松林碳收支特征[J]. 北京林业大学学报, 29(1): 1-6. |
张一平, 沙丽清, 于贵瑞, 等. 2006. 热带季雨林碳通量年变化特征及影响因子初探[J]. 中国科学D辑:地球科学, 36(增刊Ⅰ): 139-152. |
Arain A M, Restrepo-Coupe N. 2005. Net ecosystem production in a temperate pine plantation in southeastern Canada[J]. Agricultural and Forest Meteorology, 128: 223-241. DOI:10.1016/j.agrformet.2004.10.003 |
Arneth A, Kelliher F M, McSeveny T M, et al. 1998. Net ecosystem productivity, net primary productivity and ecosystem carbon sequestration in a Pinus radiation plantation subject to soil water deficit[J]. Tree Physiology, 18: 785-793. DOI:10.1093/treephys/18.12.785 |
Aubinet M, Grelle A, Ibrom A, et al. 2000. Estimates of the annual net carbon and water exchange of forest: the EUROPFLUX methodology[J]. Advances in Ecological Research, 30: 113-175. |
Baldocchi D D, Valentini R, Running S, et al. 1996. Strategies for measuring and modeling of carbon dioxide and water vapor over terrestrial ecosystems[J]. Global Change Biology, 2: 159-168. DOI:10.1111/gcb.1996.2.issue-3 |
Baldocchi D D. 2003. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future[J]. Global Change Biology, 9: 479-492. DOI:10.1046/j.1365-2486.2003.00629.x |
Fang J Y, Chen A P, Peng C H, et al. 2001. Changes in forest biomass carbon storage in China between 1949 and 1998[J]. Science, 292: 2320-2322. DOI:10.1126/science.1058629 |
Falge E, Baldocchi D, Olson R, et al. 2001. Gap filling strategies for defensible annual sums of net ecosystem exchange[J]. Agricultural and Forest Meteorology, 107: 43-69. DOI:10.1016/S0168-1923(00)00225-2 |
Greco S, Baldocchi D D. 1996. Seasonal variation of CO2 and water vapour exchange rate over a temperate deciduous forest[J]. Global Change Biology, 2: 183-197. DOI:10.1111/gcb.1996.2.issue-3 |
Guan D X, Wu J B, Zhao X S, et al. 2006. CO2 fluxes over an old, temperate mixed forest in northeastern China[J]. Agricultural and Forest Meteorology, 137: 138-149. DOI:10.1016/j.agrformet.2006.02.003 |
Hirata R, Hirano T, Saigusa N, et al. 2007. Seasonal and interannual variations in carbon dioxide exchange of a temperate larch forest[J]. Agricultural and Forest Meteorology, 147: 110-124. DOI:10.1016/j.agrformet.2007.07.005 |
Hollinger D Y, Kelliher F M, Byers J N, et al. 1994. Carbon dioxide exchange between an undisturbed old-growth temperate forest and the atmosphere[J]. Ecology, 75: 134-150. DOI:10.2307/1939390 |
Law B E, Falge E, Gu L, et al. 2002. Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation[J]. Agricultural and Forest Meteorology, 113: 97-120. DOI:10.1016/S0168-1923(02)00104-1 |
Oechel W C, Vourlitis G L, Hastings S J, et al. 2000. Acclimation of ecosystem CO2 exchange in the Alaska Arctic in response to decadal warming[J]. Nature, 406: 978-981. DOI:10.1038/35023137 |
Saigusa N, Yamamoto S, Murayama S, et al. 2005. Inter-annual variability of carbon budget components in an AsiaFlux forest site estimated by long-term flux measurements[J]. Agricultural and Forest Meteorology, 134: 4-16. DOI:10.1016/j.agrformet.2005.08.016 |
Takanashi S, Kosugi Y, Tanaka Y, et al. 2005. CO2 exchange in a temperate Japanese cypress forest compared with that in a cool-temperate deciduous broad-leaved forest[J]. Ecology Research, 20: 313-324. DOI:10.1007/s11284-005-0047-8 |
Wang H M, Saigusa N, Zu Y G, et al. 2008. Carbon fluxes and their response to environmental variables in a Dahurian larch forest ecosystem in northeast China[J]. Journal of Forestry Research, 19(1): 1-10. DOI:10.1007/s11676-008-0001-z |
Webb E K, Pearman G, Leuning R. 1980. Correction of flux measurements for density effects due to heat and water vapor transfer[J]. Quarterly Journal of the Royal Meteorological Society, 106: 85-100. DOI:10.1002/(ISSN)1477-870X |
Wilczak J M, Oncley S P, Stage S A. 2001. Sonic anemometer tilt correction algorithms[J]. Boundary Layer Meteorology, 99: 127-150. DOI:10.1023/A:1018966204465 |
Yu G R, Zhang L M, Sun X M, et al. 2008. Environmental controls over carbon exchange of three forest ecosystems in eastern China[J]. Global Change Biology, 4: 2555-2571. |
Zhang J H, Han S J, Yu G R. 2006. Seasonal variation in carbon dioxide exchange over a 200-year-old Chinese broad-leaved Korean pine mixed forest[J]. Agricultural and Forest Meteorology, 137: 150-165. DOI:10.1016/j.agrformet.2006.02.004 |