 2010, Vol. 46
2010, Vol. 46文章信息
- 陈俊华, 刘兴良, 何飞, 刘世荣
- Chen Junhua, Liu Xingliang, He Fei, Liu Shirong
- 卧龙巴朗山川滇高山栎灌丛主要木本植物种群生态位特征
- Niche Characteristics of Dominant Woody Populations in Quercus aquifoliodes Shrub Community in Balangshan Mountain in Wolong Nature Reserve
- 林业科学, 2010, 46(3): 23-28.
- Scientia Silvae Sinicae, 2010, 46(3): 23-28.
-
文章历史
- 收稿日期:2009-01-12
-
作者相关文章



2. 四川省林业科学研究院 成都 610081;
3. 中国林业科学研究院 北京 100091
2. Sichuan Academy of Forestry Chengdu 610081;
3. Chinese Academy of Forestry Beijing 100091
自Grinnell(1917)提出生态位概念以来,生态位研究成了生态学的一个热点领域。20世纪90年代,生态位这一概念在生态学界受到前所未有的关注(Leibold, 1995)。生态位理论在种间关系、群落结构、物种多样性及种群进化研究中已被广泛应用(Harper, 1977; 苏志尧等, 2003),对生态位的研究与应用已成为现代生态学的核心内容之一(尚玉昌, 1988; 钟章成, 1988)。
川滇高山栎(Quercus aquifoliodes),又称巴朗栎,喜光、耐寒、抗风且耐干旱瘠薄土壤,特产于横断山区海拔2 000~4 000 m的地带,形成了我国西部独特的高山硬叶常绿阔叶林。在分布区内,当纬度或海拔较低时,川滇高山栎多为高大乔木,而当纬度或海拔较高时,则多为矮小灌丛。川滇高山栎灌丛根系发达,萌生能力强,具有良好的水土保持和水源涵养作用。近年来,国内专家和学者对川滇高山栎作过生物量(刘兴良, 2006)、种-面积关系以及遗传方面(李进等, 1997; 1998)的研究,但对其群落生态位的研究未见报道。开展川滇高山栎灌丛主要木本植物种群生态位的研究,将有助于了解川滇高山栎群落各物种的地位与作用,了解其他物种与川滇高山栎的相互关系,为进一步研究川滇高山栎灌丛的群落稳定性、演替和天然林种间关系提供理论基础, 为正确评价与合理经营川滇高山栎灌丛提供技术支撑。
1 研究区概况研究区位于四川卧龙自然保护区皮条河上游巴朗山的阳坡(102°52′—103°24′E, 30°45′—31°25′N)。气候干湿季分明。据该区卧龙亚高山暗针叶林生态系统定位研究站(海拔2 700 m处)无林地气象观测资料表明:该区年平均气温5.3 ℃,2月最低气温-14.9 ℃,7月最高气温26 ℃,年降水量711 mm,年蒸发量827 mm,年均相对湿度79%,年日照时数1 421 h。林下土壤主要为山地棕褐土,较贫瘠、干燥,石砾含量多。川滇高山栎灌丛集中分布于巴朗山海拔2 400~3 600 m处。该群落灌木层主要植物有川滇高山栎、平枝栒子(Cotoneaster dielsianus)、木帚栒子(Cotoneaster dielsianus)、牛头柳(Salix dissa)、甘肃瑞香(Daphne tangutica)和鞘柄菝葜(Smilax stans)等;草本层主要植物有光柄糙野青茅(Deyeuxia levipes)、高山冷蕨(Cystopteris Montana)和小花金挖耳(Carpesium minus)等。
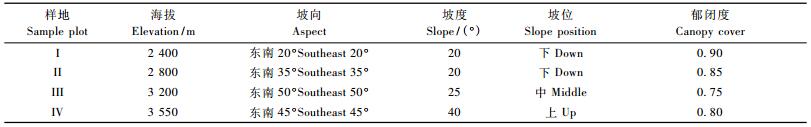
2 研究方法 2.1 样地调查在川滇高山栎灌丛分布地带进行全面线路踏查,在踏查的基础上,分别在2 400,2 800,3 200和3 550 m 4个海拔梯度各设1个25 m×25 m的典型样地,样地概况见表 1。将每个样地划分为25个5 m×5 m的小样方,记录每个样方内的木本植物种类、株数或丛数、高度、冠幅和地径等指标。
|
|
乔木层重要值=(相对密度+相对频度+相对高度)/3。
生态位宽度采用Levins(1968)和Hurlbert(1978)的生态位宽度公式计算。Levins生态位宽度Bsw计算公式为
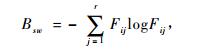
|
(1) |
式中: Bsw为种i的Levins生态位宽度; Fij为种i对第j个资源的利用占它对全部资源利用的频度,即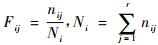
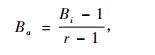
|
(2) |
式中: 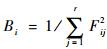
生态位相似比例指2个物种利用资源的相似性程度,其计算公式(吴刚等, 1999)为
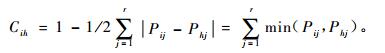
|
(3) |
式中: Cih为物种i与物种h的生态位相似性比例值,显然Cih=Chi,具有值域[0, 1];Pij和Phj分别为物种i和物种h在资源位j上的重要值。
生态位重叠指一定资源序列上,2个物种利用同等级资源且相互重叠的情况。生态位重叠值采用Levins生态位重叠公式计算(吴刚等, 1999; 史作民等, 1999; 吴大荣, 2001; 余树全等, 2003; 赵永华等, 2004; 冶民生等, 2006)。其计算公式为:
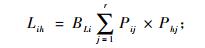
|
(4) |
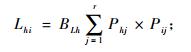
|
(5) |
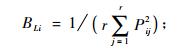
|
(6) |
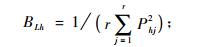
|
(7) |
式中: Lih为物种i与物种h的生态位重叠指数, Lhi为物种h与物种i的生态位重叠指数, Lih和Lhi具有域值[0, 1], BLi和BLh分别为物种i物种h的Levins生态位宽度指数, 它们都具有域值[1/r, 1]。
将原始调查数据整理后,输入计算机建立数据库,在Microsoft Visual FoxPro. 6.0中编制程序计算各种群的重要值,在Excel 2003中计算生态位宽度及生态位重叠指数。
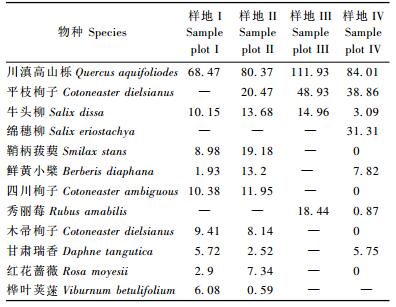
3 结果与分析 3.1 主要木本植物重要值根据川西山地巴朗山4个梯度川滇高山栎群落的样地资料计算了主要木本植物重要值(表 2)。结果表明, 在每一个资源位(海拔梯度)中,川滇高山栎种群都占绝对优势,4个资源位中川滇高山栎重要值之和为344.78,占所有种群重要值之和的53.0%。在4个资源位中,川滇高山栎矮林在海拔3 200 m梯度处重要值最大,占整个资源位重要值之和的32.46%。此外,平枝栒子种群也具有较大的优势,其重要值之和占全部种群重要值的16.64%。而其他种群则占较小的优势,且重要值之和都相差不大。
|
|
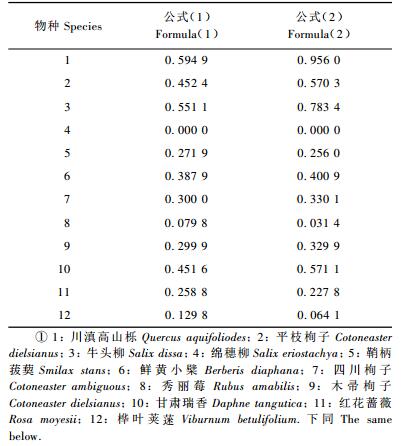
生态位宽度是度量植物种群对资源环境利用状况的尺度,种群生态位宽度越大,它对环境的适应能力越强,对资源的利用越充分。用Levins和Hurlbert2种计测生态位宽度的公式计算川滇高山栎灌丛主要木本植物的生态位宽度,其结果基本一致(表 3)。在所有种中,川滇高山栎和牛头柳的生态位宽度较大,在每个资源位(海拔梯度)中均有出现。表明川滇高山栎、牛头柳在林内分布范围较广、数量较多、利用资源较为充分,对所在环境具有较强的适应能力。12个物种的Levins生态位宽度大多集中在0.2~0.6之间,占75.0%。
|
|
川滇高山栎灌丛主要木本植物的生态位相似性比例计算结果表明(表 4),生态位相似性比例值在0.5以上的共有26对,占全部对数的39.39%,这说明川滇高山栎主要木本植物种群之间的相似性比例值较大。川滇高山栎种群与其他种群的相似性比例均较大,11对相似性比例值中大于0.4的就有8对,大于0.5的有4对:与平枝栒子、牛头柳、鲜黄小檗(Berberis diaphana)和甘肃瑞香的相似性比例值分别为0.757,0.830,0.561和0.622,尤其是跟牛头柳的相似性比例达到了0.830。从生态位相似性比例分配格局可见, 大部分种群的生态位相似性比例值为0~0.7,其中0.4~0.6的最多,共占全部种对的36.37%。
|
|
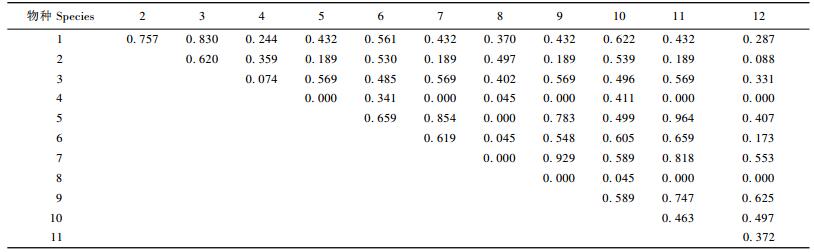
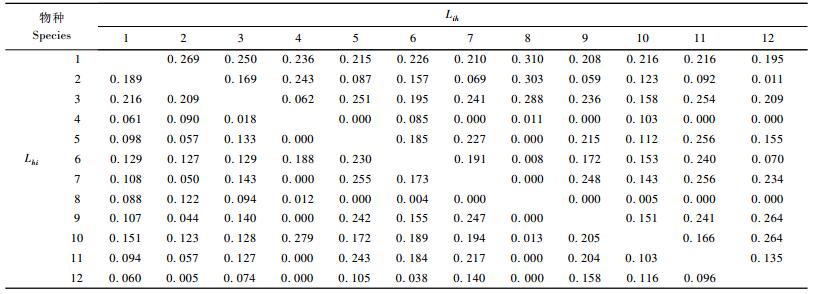
川滇高山栎灌丛主要木本植物生态位重叠计算结果表明(表 5),具有生态位重叠的种对数共有112对,占总对数的84.85%。有82对的重叠值>0.1,约占62.12%,有39对的重叠值>0.2,约占29.55%。生态位宽度大的种群之间一般能产生较大的重叠值,如川滇高山栎与平枝栒子、牛头柳的Lih重叠值分别为0.269和0.250。生态位宽度大的种群与生态位宽度小的种群也能产生较大的重叠值,如川滇高山栎与秀丽莓的Lih重叠值为0.310,甚至比与生态位宽度大的种之间的重叠值还大。而生态位宽度小的种群一般不会与生态位宽度大的种群间产生较大的重叠值,如绵穗柳(Salix eriostachya)与其他种群间的Lih生态位重叠值均不超过0.1。从生态位重叠分配格局来看,主要木本植物的Lih重叠值集中在0~0.32,Lhi重叠值集中在0~0.28,大于0.08的Lih和Lhi重叠值分别占72.74%和65.16%,表明各种群对资源的共享趋势较为明显,显示川滇高山栎灌丛群落相对稳定。
|
|
分别将卧龙巴朗山川滇高山栎灌丛主要木本植物的生态位宽度与重要值变异系数进行线性回归。结果表明(表 6),Bsw与Ba均与重要值变异系数呈典型负相关,其相关系数分别为+0.985 6和+0.973 7。表明各物种在各资源位中分布的重要值变异系数越小,其生态位宽度越大。
|
|

通常物种生态位宽度的大小决定于其对资源的利用和对环境的适应能力。重要值之和大且在资源位中分布频度大生态位宽度也较大,而重要值之和大但分布频度小的种群生态位宽度不一定大,例如牛头柳在4个资源位中均有分布,而平枝栒子只在海拔2 800,3 200和3 600 m这3个资源位中分布,平枝栒子的重要之和(108.26)比牛头柳(41.88)大2倍多,而平枝栒子的Bsw值(0.452 1)却比牛头柳(0.551 1)小。种群的生态度宽度还与重要值在资源位中分布的均匀程度有关,重要值之和大且在各资源位中分布均匀,其生态位宽度大。如秀丽莓和木帚栒子均分布于2个资源位中,但前者在海拔3 200和3 500 m资源位中的重要值分别为18.44和0.87,相差非常大,而后者在海拔2 400和2 800 m的重要值分别为9.41和8.14,差别较小。秀丽莓的重要值之和(19.31)比木帚栒子(17.55)大,但BSW值(0.079 8)却比木帚栒子(0.299 9)小。还可以看出,只在1个资源位中分布的种群,无论重要值之和多大,生态位宽度都最小,Bsw和Ba都为0。
生态位宽度与生态位相似性比例有一定关系,一般生态位宽度大的种群与其他种群间的相似性比例较大。生态位宽度小的种群与其他种群之间的相似性比例较小,如绵穗柳和秀丽莓的生态位宽度较小,它们跟其他种群间的相似性比例值均没有大于0.5的(表 4)。
生态位宽度的测定方法较多(王刚等, 1984;李意德, 1994;杨远兵等, 2001;张远东等, 2001;郑元润, 1999;刘家珍等, 2004;沈泽昊等, 2001)。但计测方法都是基于单一资源轴的,随着多维生态位计测方法的不断进步以及多元统计分析方法在其中所扮演的角色越来越重要,今后多维生态位研究将会得到进一步的发展。此外,今后计算各个种群的生态位宽度和生态位重叠指标将会采用多种指标的综合分析和比较,而不仅限于目前样地的个体数、胸高断面积、生物量和重要值等指标。
李进, 陈可咏, 李渤生. 1998. 不同海拔高度川滇高山栎群体遗传多样性的变化[J]. 植物学报, 40(8): 761-767. |
李进, 陈可咏, 李渤生. 1997. 川滇高山栎群体遗传结构的初步研究[J]. 北京林业大学学报, 19(2): 93-98. |
李意德. 1994. 海南岛尖峰岭热带山地雨林主要种群生态位特征研究[J]. 林业科学研究, 71(1): 78-85. |
刘家珍, 陈亚宁, 张元明. 2004. 塔里木河中游植物种群在四种环境梯度上的生态位特征[J]. 应用生态学报, 15(4): 549-555. |
刘兴良. 2006. 川西巴郎山川滇高山栎林群落生态学的研究. 北京林业大学博士学位论文.
|
刘兴良, 刘世荣, 宿以明, 等. 2006. 巴郎山川滇高山栎灌丛地上生物量及其对海拔梯度的响应[J]. 林业科学, 42(2): 1-7. |
刘兴良, 岳永杰, 郑绍伟, 等. 2005. 川滇高山栎种群统计特征的海拔梯度变化[J]. 四川林业科技, 26(4): 9-15. |
沈泽昊, 方精云. 2001. 基地种群分布地形格局的两种水青冈生态位比较研究[J]. 植物生态学报, 25(4): 392-398. |
史作民, 程瑞梅, 刘世荣. 1999. 宝天曼落叶阔叶林种群生态位特征[J]. 应用生态学报, 10(3): 265-269. |
尚玉昌. 1988. 现代生态学中的生态位理论[J]. 生态学进展, 5(2): 77-84. |
苏志尧, 吴大荣, 陈北光. 2003. 粤北天然林优势种群生态位研究[J]. 应用生态学报, 14(1): 25-29. |
王刚, 赵松岭, 张鹏云, 等. 1984. 关于生态位定义的探讨及生态位重叠计测公式改进的研究[J]. 生态学报, 4(2): 119-127. |
吴大荣. 2001. 福建罗卜岩闽楠(Phoebe bournei)林中优势树种生态位研究[J]. 生态学报, 21(5): 851-855. |
吴刚, 梁秀英, 张旭东, 等. 1999. 长白山红松阔叶林主要树种高度生态位的研究[J]. 应用生态学报, 10(3): 262-264. |
杨利民, 周广胜, 杨国宏. 2001. 草地群落物种多样维持机制的研究[J]. 植物生态学报, 25(5): 634-638. |
杨远兵, 刘玉成, 钟单成. 2001. 重庆缙云山不同龄级植物种群生态位宽度研究[J]. 西南师范大学学报, 26(1): 51-26. |
冶民生, 关文彬, 吴斌, 等. 2006. 岷江干旱河谷主要灌木种群生态位研究[J]. 北京林业大学学报, 28(1): 7-13. |
余树全, 李翠环. 2003. 千岛湖水源涵养林优势树种生态位研究[J]. 北京林业大学学报, 25(2): 18-23. |
赵永华, 雷瑞德, 何兴元, 等. 2004. 秦岭锐齿栎林种群生态位特征研究[J]. 应用生态学报, 15(6): 913-918. |
张远东, 潘晓玲, 顾峰雪, 等. 2001. 阜康荒漠植被灌木与半灌木种群生态位的研究[J]. 植物生态学报, 25(6): 741-745. |
郑元润. 1999. 大青沟森林植物群落主要木本植物生态位研究[J]. 植物生态学报, 23(5): 475-479. |
钟章成. 1988. 常绿阔叶林生态学研究[M]. 重庆: 西南师范大学出版社: 90-102.
|
Grinnell J. 1917. The niche-relationship of the Califomia Thrashe[J]. Auk, 34: 427-433. DOI:10.2307/4072271 |
Harper J L. 1977. Population biology of plant[M]. New York: Academic Press.
|
Hurlbert S H. 1978. The measurement of niche overlap and some relatives[J]. Ecology, 59(1): 66-77. |
Leibold M A. 1995. The niche concept revisited: mechanisitic models and community context[J]. Ecology, 76(5): 1371-1382. DOI:10.2307/1938141 |
Levins R. 1968. Evolution changing environment: some theoretical exploeations[M]. Princetion: Princetion University Preess.
|