 2007, Vol. 43
2007, Vol. 43文章信息
- 田大伦, 付晓萍, 方晰, 项文化.
- Tian Dalun, Fu Xiaoping, Fang Xi, Xiang Wenhua.
- 模拟酸雨对樟树幼苗光合特性的影响
- Effect of Simulated Acid Rain on Photosynthetic Characteristics in Cinnamomum camphora Seedlings
- 林业科学, 2007, 43(8): 29-35.
- Scientia Silvae Sinicae, 2007, 43(8): 29-35.
-
文章历史
- 收稿日期:2006-07-19
-
作者相关文章


近十几年来,随着工业的发展,SO2和NOx的排放量日渐增多,酸雨的问题越来越突出,同时酸雨危害正日益受到人们的关注,它和“温室效应”、“臭氧层破坏”一起被认为是人类面临的三大灾难性事件。我国酸雨正呈蔓延之势,是继欧洲、北美之后世界第三大重酸雨区。20世纪90年代中期,我国酸雨区已扩大到长江以南、青藏高原以东及四川盆地的广大地区,面积扩大了100多km2。以长沙、赣州、南昌、怀化为代表的华中酸雨区现已成为我国酸雨污染最严重的地区,其中心区年降酸雨频率高于90%,几乎到了逢雨必酸的程度。酸雨对森林植物的危害表现为多方面:破坏植物叶片结构(Hindawi,1980)、降低叶绿素含量(Ferenbangh,1976;吴杏春等,2004)、影响叶绿素荧光特性(马博英等,2006)、增大植物叶片膜的透性(周青等,2002)、影响植物酶活性(齐泽民等,2001;吴杏春等,2004)和光合放氧(李庆新等,1993)等。樟树(Cinnamomum camphora)是我国亚热带常绿阔叶林的主要树种,也是珍贵树种之一,具有四季常绿、树形优美、灭菌驱虫和挥发香气等特点,也是天然樟脑、芳香油、油脂的重要资源,成为我国亚热带地区植被恢复和城镇园林绿化的重要树种(郝日明等,1989)。1985年11月30日长沙市八届人大常委会第十四次会议通过决议,确定樟树为长沙市市树,在长沙市区及周围地区广泛栽培种植,长势良好。然而关于酸雨对樟树生理特性的影响研究报道很少,仅樊后保等(1996)研究了模拟酸雨对樟树幼苗的叶片和生物量的影响。本文以樟树2年生幼苗为试验材料,比较分析不同pH值酸雨对樟树幼苗光合生理特性的影响,旨在揭示樟树对酸雨的响应机制,也可为城市绿化树种的筛选和科学管理提供依据。
1 材料与方法 1.1 试验地概况试验地设在湖南省长沙市中南林业科技大学城市生态站酸雨实验室内,酸雨实验室系不锈钢微框架结构的温室,面积22 m×6 m。实验室地处东经112°48′,北纬28°03′。当地年平均气温17.2 ℃,极端最高气温40.6 ℃,最低气温-12 ℃,年平均降雨量1 400 mm。无霜期为270~300 d,日照时数年均1 677.1 h,属典型的亚热带湿润季风气候。室内环境条件因开窗对流基本与外界环境条件保持一致。
1.2 试验材料试验苗木为2年生的樟树扦插苗,苗高为50~70 cm,于2004年10月移栽到高35 cm、直径30 cm的塑料盆中,用红壤和肥料土按7:1的体积比混合而成的土壤进行栽培。在缓苗期间,用自来水喷施,长势良好。2005年4月开始进行人工酸雨喷施。
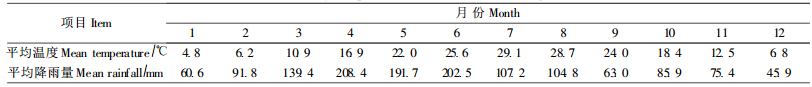
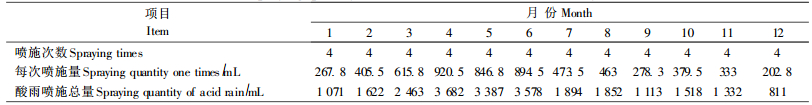
1.3 试验处理酸雨的配制按照长沙市1992—2001年酸雨中SO42-/NO3-(8:1)(蒋益民,2003),用浓硫酸和浓硝酸配制酸原液,再用蒸馏水稀释pH值为3.0、4.0、5.0的酸雨原液,同时进行对照喷施试验,每组试验设4个重复。以长沙市1971—2000年每月平均降雨量(表 1)为依据,按盆口总面积,折算成每个pH值等级人工浇灌苗木酸雨量(表 2),每次喷洒至叶片彻底湿润,对照组苗木用pH7.0的自来水等量喷施。
|
|
|
|
于2005年6月中旬,选择晴朗天气,从9:00—11:00进行,测定时保持外界正常的温度、湿度和CO2浓度,利用系统红蓝光源设置2 500,2 000,1 500,1 100,1 000,500,350,200,100,50,30,10,0 μmol·m-2s-1光合有效辐射,每个点最短等待时间是180 s,最长等待时间是240 s,仪器标定好进入工作状态后,自动记录测定数据。
1.4.2 光合日进程采用Licor-6400p光合测定系统对樟树幼苗叶片进行连体光合测定。测定时间为2005年6月下旬,选择晴朗天气,从8:00—18:00,每间隔2 h测定1次,重复测定3 d。在每株樟树上部选择处理后新萌发的当年生成熟叶片3片,每片重复记录5组数据,取其平均值。测定的指标有:净光合速率Pn(μmol CO2·m-2s-1)、蒸腾速率Tr(mmolH2O·m-2s-1)、光合有效辐射PAR(μmol·m-2s-1)、叶面大气蒸气压亏缺VPDleaf(kPa)、胞间CO2浓度Ci(μmolCO2·mol-1)、气孔导度Gs (molH2O·m-2s-1)、叶片温度tleaf (℃)、参比室CO2浓度CR(μmolCO2·mol-1)、样本室CO2浓度CS(μmolCO2·mol-1)、参比室H2O浓度CH2OR(mmolH2O·mol-1)、样本室H2O浓度CH2OS(mmolH2O·mol-1)、样本室内相对湿度RHS (%)、参比室内相对湿度RHR(%)、冷却器温度tBlk(℃)、样本室温度tair(℃)。这些指标在仪器上同步显示。
1.4.3 日均净光合速率季节变化在2005年6月、9月、12月和2006年3月下旬分别选择3个晴朗的天气,测定不同酸雨处理樟树的净光合速率,从8:00—18:00,每间隔2 h测定1次,取6个测定时间的平均值为日平均净光合速率。
1.5 数据处理采用低辐射强度下(200 μmol·m-2s-1以下)光合有效辐射与净光合速率相关方程,计算表观量子效率(AQY,molCO2·mol-1 photon)、暗呼吸速率(Rd,μmol·m-2s-1)和光补偿点(LCP,μmol·m-2s-1)。AQY用净光合速率对光合有效辐射响应曲线的起始斜率来表示(潘晓云等,2002;朱万泽等,2001)。光饱和点为曲线中净光合速率最高时的光合有效辐射。
气孔限制值(Ls)按简化公式Ls=(Ca-Ci)/(Ca-J)求得。式中:Ca是大气CO2浓度;Ci是胞间CO2浓度;J是CO2补偿点,忽略不计。
采用Excell软件和SPSS13.0软件进行数据处理。
2 结果与分析 2.1 不同pH值酸雨处理樟树苗木的光合作用-光响应如图 1所示,不同pH值酸雨对樟树的光合作用-光响应曲线影响不同。当PAR低于200 μmol·m-2s-1时,不同处理的樟树Pn均与PAR呈线性关系,且随着PAR强度的增强而增大,同时不同处理间的Pn差别不大;当PAR在200~2 500 μmol·m-2s-1之间,Pn随光强的增加而缓慢增大,不同酸雨处理间的Pn增大;同一光合有效辐射强度下,pH3.0、4.0酸雨处理的樟树Pn均明显高于对照,而且pH3.0酸雨处理的Pn高于pH4.0处理的Pn,pH5.0酸雨处理的Pn低于对照。这说明pH3.0、pH4.0酸雨增强了樟树利用200 μmol·m-2s-1以上的光合有效辐射的能力。

|
图 1 不同pH值酸雨处理的樟树光合作用-光响应曲线 Fig. 1 The curve of Pn-PAR of C. camphora under different acid rain treatments |
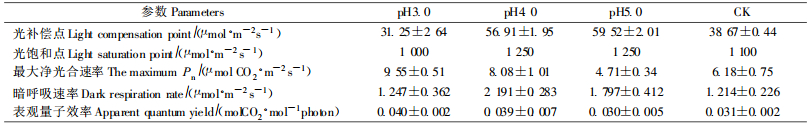
由表 3可以看出,不同pH值酸雨对樟树的光补偿点、光饱和点、最大净光合速率、暗呼吸速率、表观量子效率均产生了明显的影响。与对照相比,pH3.0酸雨处理后,樟树的光补偿点、光饱和点分别下降19.19%、9.09%,而pH4.0、5.0酸雨处理后,光补偿点分别增高47.17%、53.92%,光饱和点均增高13.64%;pH3.0、4.0酸雨处理增高樟树最大净光合速率和表观量子效率,其中最大净光合速率分别增高54.53%、30.74%,表观量子效率分别增高27.07%、22.61%,而pH5.0酸雨处理的分别下降23.79%和4.14%;pH3.0、4.0和5.0酸雨处理增高樟树的暗呼吸速率,分别增高2.70%、80.45%、47.55%。
|
|
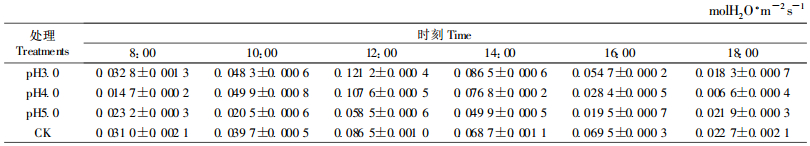
由表 4可以看出,不同pH值酸雨处理的樟树气孔导度变化也有所不同,气孔导度(Gs)最大值和日变化幅度的大小顺序是pH3.0>pH4.0>pH5.0。其中:pH3.0、4.0处理的Gs最大值分别为0.121 2、0.107 6 molH2O·m-2s-1,比对照(0.086 5 molH2O·m-2s-1)提高40.12%、24.39%,日变化幅度分别为0.102 9和0.101 molH2O·m-2s-1,较对照(0.063 8 molH2O·m-2s-1)增加了61.29%、58.31%;pH5.0的Gs最大值为0.058 5 molH2O·m-2s-1,比对照降低32.37%,日变化幅度为0.039 molH2O·m-2s-1,较对照低38.87%;Gs最小值的大小顺序是pH5.0>pH3.0>pH4.0,依次为0.019 5、0.018 3和0.006 6 molH2O·m-2s-1,较对照(0.022 7 molH2O·m-2s-1)分别降低了14.10%、19.38%、70.93%。
|
|
从表 5可知,不同pH值酸雨处理的Ci最大值的大小顺序是pH4.0>pH3.0>pH5.0,其中pH3.0、4.0处理的分别是351.32和362.02 μmolCO2·mol-1,较对照(329.7 μmolCO2·mol-1)增高6.56%、9.80%,pH5.0处理的是319.0 μmolCO2·mol-1,较对照低3.25%;Ci最小值的大小顺序是pH5.0>pH4.0>pH3.0,依次为236.4、229.4和215.62 μmolCO2·mol-1,较对照(208.8 μmolCO2·mol-1)分别增高13.22%、9.87%、3.27%;Ci日变化幅度的大小顺序是pH4.0>pH3.0>pH5.0,pH4.0处理的为132.62 μmolCO2·mol-1,较对照(120.9 μmolCO2·mol-1)增高9.69%,pH3.0、5.0处理的分别为114.92和103.38 μmolCO2·mol-1,较对照分别降低4.95%、14.49%。可见,3种酸雨均增大了樟树Ci最小值,pH4.0增大了Ci最大值和日变化幅度,pH3.0、pH5.0降低了Ci最大值和日变化幅度。
|
|
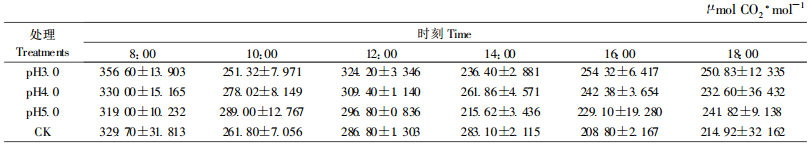
从表 6可知,不同酸雨处理的Ls最大值的大小顺序是pH5.0 >pH4.0>pH3.0,分别较对照(0.432 4)降低30.09%、26.34%、20.93%;最小值的大小顺序是pH5.0>pH3.0>pH4.0,分别较对照(0.030 9)增高360.19%、114.56%、34.95%;Ls日变化幅度的大小顺序是pH4.0>pH3.0>pH5.0,较对照(0.401 5)分别降低了33.15%、43.31%、52.35%。表明3种酸雨降低了樟树幼苗Ls的最大值和日变化幅度,增高了Ls最小值。
|
|
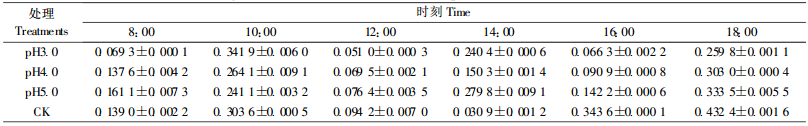
从表 7中可以看出,樟树净光合速率的日变化呈不对称的“双峰型”变化,出现峰值时间为10:00和14:00,12:00出现“光合午休”,14:00后净光合速率不断降低,直至18:00净光合速率为负值。这与“樟树幼树光合特性及其对CO2浓度和温度升高响应”的研究结果(田大伦等,2004)一致。
|
|
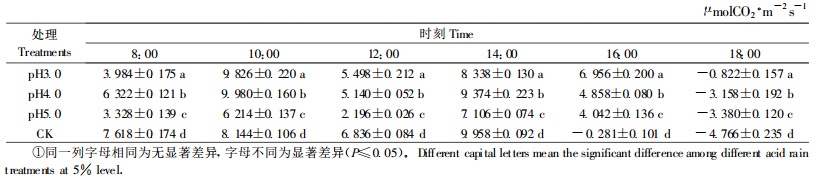
不同pH酸雨对樟树的净光合速率(Pn)日变化均有一定的影响,其影响程度因酸雨pH值的不同而有所差异。8:00,pH3.0、4.0、5.0的Pn分别比对照降低47.70%、17.01%、56.32%,并随着太阳辐射的增强和温度的升高而增大;10:00出现峰值,pH3.0、4.0的Pn分别比对照增高20.65%、22.54%,而pH5.0的Pn比对照降低23.70%;12:00,出现“光合午休”,此时pH3.0、4.0、5.0的Pn较10:00分别下降了4.328、4.84和4.018 μmolCO2·m-2s-1,降幅分别是对照(1.308 μmolCO2·m-2s-1)的3.308 9、3.700 3和3.071 9倍;14:00,出现光合第2个高峰,pH3.0、4.0的Pn较第1个峰值分别下降了15.14%和6.07%,而pH5.0的Pn较第1峰值增高了14.35%;16:00,酸雨处理的樟树Pn均有所降低,但仍为正值,而对照的Pn为负值;18:00,酸雨处理的樟树Pn降至负值,但仍高于对照。方差分析和多重比较的结果表明,同一时刻,不同pH酸雨处理的樟树Pn间差异显著(P≤0.05)。
从表 7看出,pH3.0、4.0、5.0的Pn日变化幅度分别为10.648、13.138和10.486 μmolCO2·m-2s-1,比对照(14.724 μmolCO2·m-2s-1)降低了27.68%、10.77%、28.90%。说明不同pH酸雨均增大了樟树“午休”时Pn的下降幅度,降低了Pn的日变化幅度。
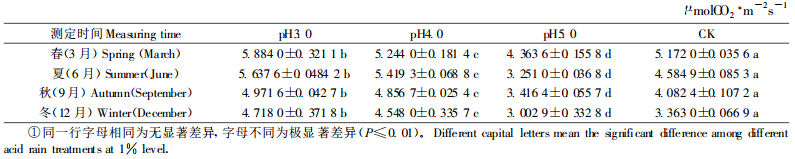
2.4 不同pH值酸雨处理的樟树幼苗日均净光合速率(Pn)的季节变化从表 8可知,酸雨对樟树日均Pn有一定的影响。同一季节不同处理的日均Pn大小顺序是pH3.0>pH4.0>pH5.0,在不同季节,pH3.0、4.0的日均Pn均高于对照,而pH5.0的日均Pn低于对照。夏季pH3.0、4.0的日均Pn分别是5.637 6和5.419 3 μmolCO2·m-2s-1,比对照分别增高22.96%、18.20%,pH5.0的日均Pn为3.251 0 μmolCO2·m-2s-1,比对照低29.09%;秋季pH3.0、4.0的日均Pn比对照分别高21.78%、18.97%,pH5.0的日均Pn比对照低16.31%;冬季pH3.0、4.0的日均Pn比对照分别高40.29%、35.24%,pH5.0的日均Pn比对照低10.71%;春季pH3.0、4.0的日均Pn比对照分别高13.77%、1.39%,pH5.0的日均Pn比对照低15.63%。方差分析和多重比较结果表明,不同pH值酸雨处理间差异极显著(P≤0.01),pH5.0酸雨降低了樟树的日均Pn,pH3.0、4.0酸雨提高了樟树的日均Pn。
|
|
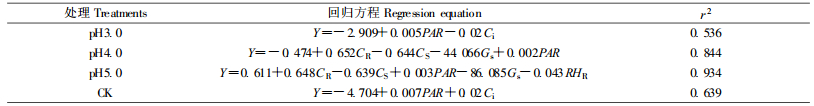
净光合速率的变化是各生理生态因子综合作用的结果,不同阶段各因子的影响不同,甚至效应相反(郭天财,2002;吴楚等,2005;冯建灿等,2002)。利用生长季节所获得的不同pH值酸雨处理的樟树Pn及其影响因子的数据,以Y代表净光合速率(Pn)对Gs、Ci、Tr、VPDleaf、tair、tleaf、tBlk、CR、CS、CH2OR、CH2OS、RHR、RHS、PAR进行多元回归分析。
从表 9可知,pH3.0酸雨处理的Pn的主要影响因子是PAR、Ci,其中与PAR呈正相关,与Ci呈负相关,且Ci对Pn的影响程度大于PAR;pH4.0的Pn的主要影响因子是CR、CS、Gs、PAR,其中与CR、PAR呈正相关,与CS、Gs呈负相关,各因子对Pn影响程度的大小次序是Gs>CR>CS>PAR;pH5.0的Pn主要影响因子是CR、CS、RHR、PAR、Gs,其中与CR、PAR呈正相关,与RHR、CS、Gs呈负相关,各因子对Pn影响程度的大小顺序是Gs>CR>CS>RHR>PAR。对照的主要影响因子是Ci、PAR,且均呈正相关,Ci对Pn的影响程度大于PAR。
|
|
pH3.0、4.0酸雨增强了樟树利用200 μmol·m-2s-1以上光合有效辐射的能力。pH3.0酸雨降低了樟树的光补偿点、光饱和点。pH3.0、4.0酸雨处理增大了樟树最大净光合速率和表观量子效率。3种酸雨均提高了樟树的暗呼吸速率,这可能是樟树叶片细胞中线粒体遭到破坏或解体,氧化磷酸化解偶联,因此暗呼吸速率增大(邱栋梁等,2002;文宗振等,1992)。
3种酸雨均降低了樟树气孔导度的最小值,pH3.0、4.0酸雨提高了樟树气孔导度的最大值和日变化幅度。酸雨均提高了樟树胞间CO2浓度最小值,降低了其气孔限制值的日变化幅度。植物气孔不仅可以防止植物因蒸腾作用所引起的水分过分损失,确保植物获得足量的CO2气体进行光合作用,而且也是调控土壤-植被-大气连续体间同水分损失和碳素获取有关的物质和能量交换的关键环节(王玉辉等,2000)。酸雨作用下樟树气孔导度最小值降低说明樟树为适应酸性条件,气孔导度下降,降低蒸腾,减少水分损失。
樟树幼苗的净光合速率日变化均呈不对称的“双峰型”变化,峰值出现时间为10:00、14:00,12:00出现“光合午休”,经不同pH值酸雨处理后,樟树的“光合午休”现象更为明显,且酸雨降低了樟树净光合速率的日变化幅度。引起叶片光合“午休”的植物自身因素有2个,一个是气孔的部分关闭,另一个是非气孔限制;根据Farquha和Sharkey(1982)的观点, 当叶内胞间CO2浓度减少,气孔限制值增大时,气孔的部分关闭成为光合下降的主要原因。而当胞间CO2浓度增加,气孔限制值减小时,非气孔限制成为光合下降的主要原因。本研究中不同酸液处理的樟树幼苗和对照的Ci值、Ls值12:00时均较10:00时高,这说明3种酸雨处理的樟树幼苗和对照的光合“午休”原因均为气孔的部分关闭。而酸雨处理的樟树光合“午休”现象较对照明显,可能是因为樟树在高强度的太阳有效辐射和酸性胁迫条件下为避免伤害而产生的适应。
本研究中,pH3.0、4.0酸雨处理的樟树日均Pn高于对照的日均Pn。而有研究结果(李庆新等,1993;蔡世舫等,2000;邱栋梁等,2002)表明,随着酸雨pH值的降低,植物受伤害的程度增大,植物的净光合速率也随着酸雨的pH值的降低而降低。而樊后保等(1996)的研究表明,pH2.0酸雨处理对樟树幼苗生长的影响是十分明显的,它增加了叶细胞膜的透性,加速了阳离子的淋失,使细胞原生质酸化,降低了叶绿素的含量,因而伤害了幼苗的叶片,抑制了生长;pH3.5酸雨处理尽管对幼苗叶片造成一定的伤害,但却促进了幼苗和地径的增长,增加了单株生物量。Wood和Bormann(1976)在对北美乔松(Pinus strobus)的模拟酸雨试验中也发现,pH4.0以下的模拟酸雨刺激了幼苗的生长,并指出是由于酸雨中NO3-的施肥作用的效果。也有研究(杨振德,1999)认为,植物的形态结构特点和生物学特性决定其对酸雨所造成的伤害的自身恢复功能和抗性,叶面光滑、蜡质层厚、叶革质、坚韧的较叶面粗糙、凹陷薄质的抗酸雨能力强,叶面多毛、能分泌粘性物质的不易受害。从樟树的叶片形态结构上看,具备叶面光滑并且有蜡质层的特点,付晓萍等(2006)在酸雨对植物形态影响的研究中发现,樟树幼苗在pH3.0、4.0、5.0的酸雨作用下根冠比增大,说明樟树在酸性条件下根系生长速度加快,吸收营养物质的能力相对增强。表明樟树耐酸性强,在酸雨区可作为绿化树种广泛栽种。
本文对3种酸雨处理的2年生樟树幼苗光合特性进行了研究,随着时间的推移和樟树年龄的增大,樟树的光合特性是否会发生变化尚需进一步研究。
蔡世舫, 任清文, 欧红梅, 等. 2000. 酸雨对小麦生理特性影响的研究. 安徽农业科学, 28(3): 303-304. DOI:10.3969/j.issn.0517-6611.2000.03.022 |
樊后保, 藏润国. 1996. 模拟酸雨对樟树种子萌发和幼苗生长的影响. 浙江林学院学报, 13(4): 412-417. |
付晓萍, 田大伦, 黄智勇. 2006. 模拟酸雨对植物形态学效应的影响. 浙江林学院学报, 23(5): 521-526. DOI:10.3969/j.issn.2095-0756.2006.05.010 |
郝日明, 魏宏图. 1999. 紫金山森林植被性质与常绿落叶阔叶混交林重建可能性的探讨. 植物生态学报, 23(2): 108-115. DOI:10.3321/j.issn:1005-264X.1999.02.002 |
蒋益民, 曾光明, 张龚, 等. 2003. 长沙市大气湿沉降化学及变化特征. 城市环境与城市生态, 16(增刊): 23-25. |
李庆新, 谢田, 徐中际. 1993. 酸雨对菠菜光合作用的影响. 环保科技, 15(2): 1-21. |
马博英, 徐礼根, 蒋德安. 2006. 模拟酸雨对假俭草叶绿素荧光特性的影响. 林业科学, 42(11): 554-560. |
潘晓云, 曹琴东, 王根轩, 等. 2002. 扁桃与桃光合作用特征的比较研究. 园艺学报, 29(5): 403-407. DOI:10.3321/j.issn:0513-353X.2002.05.001 |
齐泽民, 钟章成, 邓君. 2001. 模拟酸雨对杜仲叶氮代谢的影响. 植物生态学报, 25(5): 544-548. DOI:10.3321/j.issn:1005-264X.2001.05.007 |
邱栋梁, 刘星辉. 2002. 模拟酸雨对龙眼叶绿体活性的影响. 应用生态学报, 13(12): 1559-1562. DOI:10.3321/j.issn:1001-9332.2002.12.010 |
田大伦, 罗勇, 项文化, 等. 2004. 樟树幼树光合特性及其对CO2浓度和温度升高的响应. 林业科学, 40(5): 88-92. DOI:10.3321/j.issn:1001-7488.2004.05.014 |
王玉辉, 周广胜. 2000. 羊草叶片气孔导度对环境因子的响应模拟. 植物生态学报, 24(6): 739-743. DOI:10.3321/j.issn:1005-264X.2000.06.017 |
文宗振, 钟荣亮, 潘洵操. 1992. 酸雨对萝卜叶超微结构的危害初报. 广西农业大学学报, 11(4): 85-87. |
吴杏春, 林文雄, 洪清培, 等. 2004. 模拟酸雨对草坪若干生理指标的影响. 草业科学, 21(8): 554-560. |
杨振德. 1999. 几种园林树种对酸雨抗性的研究. 广西大学学报, 3(2): 29. |
中国树木志编委会. 1989. 中国主要树种造林技术. 北京: 中国林业出版社, 531-539.
|
周青, 黄晓华, 刘小林. 2002. 酸雨对3种木本植物的胁迫效应. 环境科学, 23(2): 42-46. |
朱万泽, 王金锡, 薛建辉, 等. 2001. 四川桤木光合生理特性研究. 西南林学院学报, 21(4): 196-204. |
Farquhar G D, Sharkey T D. 1982. Stomata conductance and photosynthesis. Ann Rev Plant Physiol, 33: 317-345. DOI:10.1146/annurev.pp.33.060182.001533 |
Ferenbangh R W. 1976. Effects of simulated acid rain on Phaseolus vulgaris L. American Tonrnal Botany, 63(3): 283-288. DOI:10.1002/j.1537-2197.1976.tb11813.x |
Hindawi I J. 1980. Response of bush bean exposed to acid rain mist. American Tournal Botany, 67: 168-173. DOI:10.1002/j.1537-2197.1980.tb07638.x |
Wood T, Bormann F H.1976.Short-term effects of a simulated acid rain upon the growth and nutrient relations of Pinus strobus L//Dochinger L S, Seliga T A. Proc Internat Symp: Acid precipitation and the forest ecosystem.Upper Darby: N E For Exp Sta, 815-826
|