 2007, Vol. 43
2007, Vol. 43文章信息
- 尹伟伦, 万雪琴, 夏新莉.
- Yin Weilun, Wan Xueqin, Xia Xinli.
- 杨树稳定碳同位素分辨率与水分利用效率和生长的关系
- Relations between Stable Carbon Isotope Discrimination and Water Use Efficiency as well as Growth for Poplar
- 林业科学, 2007, 43(8): 15-22.
- Scientia Silvae Sinicae, 2007, 43(8): 15-22.
-
文章历史
- 收稿日期:2006-04-19
-
作者相关文章


2. 四川农业大学林学院 雅安 625014
2. College of Forestry, Sichuan Agricultural University Ya'an 625014
提高植物的水分利用效率(WUE)是作物遗传改良的重要目标,目前在小麦(Triticum aestivum)、大麦(Hordeum vulgare)、水稻(Oryza sativa)、花生(Arachis hypogaea)等重要农作物上已经广泛开展(张正斌, 2003; Bacon, 2004)。WUE是一个重要的产量决定因子,提高WUE可以降低获得一定产量的需水量,提高特定土壤有效水量下的总生物产量,达到节水的目的。尽管关于WUE的遗传变异的研究可以追溯到近100年前,但与植物WUE相关的生理指标(如光合、蒸腾、气孔导度等)却很少被作为选育高WUE的直接指标。这主要是因为它们的遗传力低,与产量的关系复杂,且大规模测定比较困难(Hall et al., 1994)。稳定碳同位素分辨率(Δ13C)与植物生理过程间建立联系后,评价WUE的遗传变异获得巨大进步(Farquhar et al., 1984;1989; Udayakumar et al., 1998; Richards et al., 2002)。在干旱胁迫下,很多作物的Δ13C可以作为气孔导度和水分利用效率的可靠指标(Araus et al., 1993; Matus et al., 1996)。Δ13C与产量的关系受到环境条件、作物的物候期和用来分析Δ13C的植物器官的影响(Condon et al., 1992a; Araus et al., 1997;2003),可以表现为正相关、负相关,甚至不相关(Condon et al., 1987; Craufurd et al., 1991; Merah et al., 2002)。许多C3植物种实的Δ13C存在丰富的遗传变异,并且这种变异具有高的广义遗传力[如在硬质小麦(Triticum durum)中为0.76~0.85(Merah et al., 2002)]和低的基因型×环境互作效应(G×E)(Ismail et al., 1992; Condon et al., 1992b; Richards, 1996; Lambrides et al., 2004)。这些特点使Δ13C成为选育高WUE植物品种的一个重要指标。Δ13C是用来评价整株水平的WUE和选育节水基因型的有力工具,已应用于小麦、大麦、水稻、棉花(Gossypium hirsutum)等作物的遗传改良和育种工作(Farquhar et al., 1984; Hubick et al., 1989; Read et al., 1991; Rebetzke et al., 2002; Sheshshayee et al., 2003; Impa et al., 2005; Stiller et al., 2005; 张正斌,2003)。
杨树(Populus)是世界上栽培面积最广、数量最多的人工林树种,对经济发展、环境建设发挥着重要的作用。我国是世界上杨树人工林面积最大的国家。我国的华北和西北地区干旱少雨,水资源匮乏,这严重地制约着杨树人工林的发展。耐旱杨树品种和高水分利用效率品种的选育是杨树育种的重要方向之一,也是缓解和充分利用三北地区有限水资源的有效途径。干旱条件下杨树叶片δ13C与水分利用效率呈显著正相关,它是间接评价同等水分条件下不同杨树无性系水分利用效率高低的可靠指标(赵凤君等, 2005)。目前尚未有用Δ13C评价田间生长条件下杨树不同无性系水分利用效率高低,及Δ13C与杨树田间生长量间关系的研究报道。本研究的目的是用杨树不同器官的Δ13C来揭示大田条件下杨树在不同时间尺度上的水分利用效率变化,及Δ13C与杨树生长的关系。这对Δ13C在杨树水分利用效率评价上的应用及杨树高水分利用效率无性系选育具有重要的理论和应用价值。
1 材料与方法2004年3月从美国引进25个杨树无性系,将插条截成15 cm的插穗,在清水中浸泡24 h后,扦插于试验地中,扦插的株行距为100 cm × 75 cm,每个无性系插40个插穗。插后及时浇水、除草、抹芽,不施肥。试验地位于北京市海淀区东升乡北林科技苗圃内。试验地立地条件均一。2005年3月未萌动前进行疏伐,保留密度为200 cm × 150 cm。7月下旬从中选择杂交组合不同和表型差异大、生长良好的无性系DN182 (P. deltoides × P. nigra)、SIMPLOT [P. nigra × (P. deltoides × P. nigra)]、NM6 (P. nigra × P. maximowiczii)、311/93 (P. trichocarpa × P. nigra)、R270 (P. deltoides × P. nigra)、PC1 (P. deltoides × P. nigra)、59/289 (P. trichocarpa × P. deltoides)为研究对象,分别取样测定各无性系功能叶、当年生枝、树干的有机碳同位素组成。取样方法为:每无性系选取典型样株3株,每株取其主干顶梢第6~10个叶片形成混合叶样;取最上端的3个当年生急发枝(proleptic branching)(上年形成的侧芽,在次年萌发形成的枝条),截取与树干相接处10 cm形成混合枝样品;伐倒样株,取离地面15 cm高处的10 cm树干形成混合干样品。采集的样品及时带回实验室,叶样用蒸馏水冲洗表面3次,与去皮后的枝、干样品一起于80 ℃烘干。烘干的叶样去主叶脉后研磨成粉末,过100目筛收集分析样品;烘干的树干样品和枝样品先用锋利刀片削成细小薄片,再研磨成粉末。叶、枝、干的粉末样品用真空密闭高温氧化法获得纯净CO2,用MAT253同位素质谱仪测量其碳同位素比值,每样品进行2次独立分析。具体步骤如下:1)将5 mg粉末样品放进石英反应管内,然后放入处理好的氧化剂5 g;2)将装有样品和氧化剂的石英管焊接到真空系统上,边用电吹风加热边抽真空至真空10-2 Pa用火焰煅取下;3)将石英管放到马福炉内加热900 ℃ 5 h,冷却取出,将石英管放进套管内,接到真空系统上;4)打开高真空开关和真空系统上的套管活塞及样品管,抽取真空至10-4 Pa;5)配制冷凝液套在冷井上,关闭真空活塞,折断石英管,使其气体析出,样品管底部套上液氮杜瓦瓶吸收CO2;6) 3 min后打开高真空活塞,毛细抽走冻不住的杂质气体,得到纯净的CO2气体,关闭样品活塞,送质谱测试(样品的氧化和同位素测定均在“中国科学院地质与地球物理研究所岩石圈构造演化国家重点实验室”完成)。
δ13C、Δ13C和WUE等的计算方程(Bert et al., 1997; Hones,2004)如下:
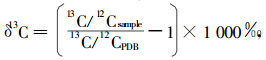
|
(1) |
式中:13C/12Csample是指植物样品中13C/12C的比值,13C/12CPDB是指标准PDB化石中13C/12C的比值。
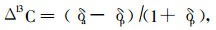
|
(2) |
式中:δa是指大气中CO2的δ13C值,这里取7.7‰,δp是植物样品的δ13C值,由方程(1)求得。
将Δ13C通过下列方程(Marshall et al., 1994)转换为水分利用效率(WUE):
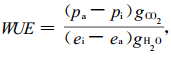
|
(3) |
式中:Pa是空气CO2偏压,Pi是胞间CO2偏压,gCO2是叶片对CO2的总导度,gH2O是叶片对水气的总导度,ei是胞间水气偏压,ea是大气水气偏压。方程(3)中的导度比用0.6代替得到方程(4)。
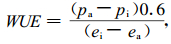
|
(4) |
式中:pa取实测平均值355 μmol·mol-1 air与标准大气压(PB, Pa)的乘积,即pa=355×PB/1 000=355×10-6×101 325=35.97 Pa。ei用方程(5)求得:
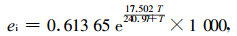
|
(5) |
式中:T值为2004年7月24日、8月17日和8月29日3天7:00—18:00叶温日进程(LI6400光合仪测定)的平均值,T=average(32.9, 32.8, 32.3)=32.7 ℃。由方程(5)求得ei=4 967.6 Pa。ea取这3天日进程大气水分的偏摩尔分压,即ea=average(22.4, 22.4, 18.2)=21 m mol·mol-1 air=(21/1 000)×101 325=2 127.8 Pa。
pi/pa由方程(6)(Hones, 2004; 张正斌, 2003)求得:
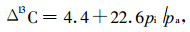
|
(6) |
将Δ13C和pa值代入方程(6),即可求得pi和(pa-pi)。
不同叶位气体交换的测定:用Li-Cor6400光合仪,于取样当日每无性系选取3株生长良好的植株,从第1片展开叶片开始,每隔4个叶片测定1个叶子的光合、蒸腾作用。仪器参数设定为:流速500 cm3·s-1,气孔比率0,叶温33 ℃,有效光合辐射(PAR)700 μmol·m-2s-1,参比CO2浓度400 μmol·L-1。气体交换日进程的测定:每个无性系随机选择3株平均木,每株依据不同叶位光合蒸腾测定的结果,在光合速率最强区域选择向南的3个叶片,作上标记,分别于7: 30、9:00、10:30、12:00、13:00、14:00、15:30、16:30、17:30定叶定位测定1次,仪器参数设定为:流速500 cm3·s-1,气孔比率0,参比CO2浓度400 μmol·L-1。数理统计分析在SPSS11.5相应模块上完成。
2 结果与分析 2.1 7个无性系瞬时水分利用效率比较杨树不同叶位叶片的水分利用效率实质上反映了叶片生长过程中的水分利用效率的动态变化(万雪琴等, 2006)。从图 1a可以看出,不同部位叶片的水分利用效率存在明显差异,这反映了叶片从发生到成熟,再到衰老过程中WUE由小变大,再逐渐减小的变化规律。从中可以看出有些无性系如SIMPLOT和PC1的叶片相对于其他叶片来说,随着叶龄的增加,WUE能在较长的时间内维持在较高的水平,从而使整株水平的WUE较高;而DN182和59/289的各部位叶片的WUE都较低,从而导致其整株水平的WUE较低。7个无性系的功能叶在典型夏日晴天条件下的WUE日变化的总体趋势是由大变小,从图 1b可以看出有些无性系(如PC1和SIMPLOT)在一天中的WUE都高于另一些无性系(如59/289和R270等)。用不同叶位叶片WUE的均值与功能叶日进程WUE均值的平均数为指标,来比较7个无性系的水分利用效率高低,所得结果为:SIMPLOT>PC1>311/93>NM6>R270>59/289>DN182(表 1)。

|
图 1 7个杨树无性系瞬时水分利用效率比较 Fig. 1 Comparison on WUE among 7 poplar clones a.不同叶位叶片的水分利用效率WUE of leaves at different positions on stem b.功能叶水分利用效率日变化Diurnal process of WUE of functional leaves |
|
|

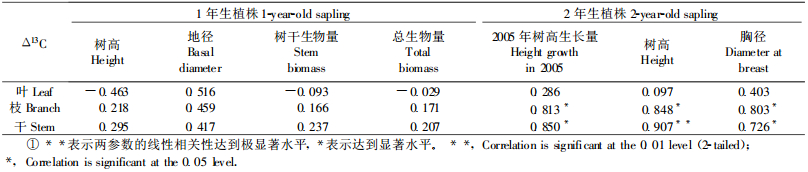
表 2是以7个无性系为样本,综合计算其叶、枝和干Δ13C值,从中可见,杨树不同器官的Δ13C差异显著,其基本规律是Δ13C功能叶>Δ13C当年生枝>Δ13C树干。由于WUE与Δ13C呈负相关,因此由Δ13C功能叶求得的WUE显著地低于枝和干的,而枝与干间的差异在95%水平不显著。对每一无性系而言,干的Δ13C显著低于枝,而WUE则高于当年生枝。植物组织碳同位素组成是由其形成过程决定的,因此叶、枝、干碳同位素组成的不同反映了不同时间尺度上WUE的差异。叶片Δ13C反映了叶片形成过程中的WUE,时间长度约为1个月(7月初到8月初);枝条Δ13C反映了从发芽(约3月底)到采样这一时间段的WUE;而干Δ13C则反映从扦插(2004年4月初)到采叶(2005年8月3日)这一时间段的平均WUE。可见,在2005年7—8月的WUE要低于4—8月时间段的,以2004年4月到2005年7月底的WUE最高。7个无性系叶、枝、干的Δ13C的样本标准差分别是0.454、0.507和0.454,说明不同无性系的当年生枝的Δ13C差异较大,故枝Δ13C相对于叶Δ13C和干Δ13C而言,对不同无性系的WUE有较高的区分能力。干Δ13C较当年生枝低,但与叶无差异,因此用干Δ13C是评价不同无性系长期WUE的较好指标。从表 2还可看出,由叶、枝和干δ13C分别求算所得的pi-pa值依次增大。由于pi-pa是CO2从大气进入气孔的推动力,故pi-pa的依次增大说明气孔摄入CO2的平均能力在叶、枝和干形成的不同时期逐渐增大。导致其增大的原因,可能是气孔对CO2的导度减少,或是光合碳同化效率的提高,或两者共同作用的结果。具体原因可以结合不同器官组织的氧同位素(18O)组成δ18O和相应的氧同位素分辨率Δ18O来确定(Bacon, 2004; Farquhar et al., 1998)。由于本研究没有测定样品的Δ18O,所以尚不能确定从叶到枝再到树干的pi-pa逐渐增加的确切原因。
|
|
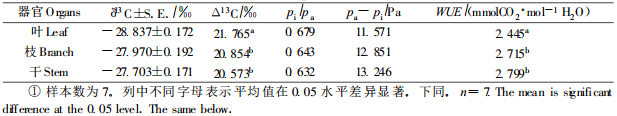
表 3列出了7个无性系功能叶片、当年生枝、树干的Δ13C及其相应的WUE值。功能叶的Δ13C和WUE除NM6和311/93差异不明显外,其他无性系均存在显著差异;而当年生枝条Δ13C和树干Δ13C及相应的WUE在7个无性系间彼此均表现出显著差异。重复样品间的δ13C离散程度低,样本标准差小(21个独立样本,2次重复的标准差平均值为0.013,最大标准差为0.049),使无性系间在Δ13C和WUE上表现出广泛的显著性差异(表 3)。这说明用Δ13C评价不同无性系的整株水平的长期WUE,具有较高的判别能力。
|
|
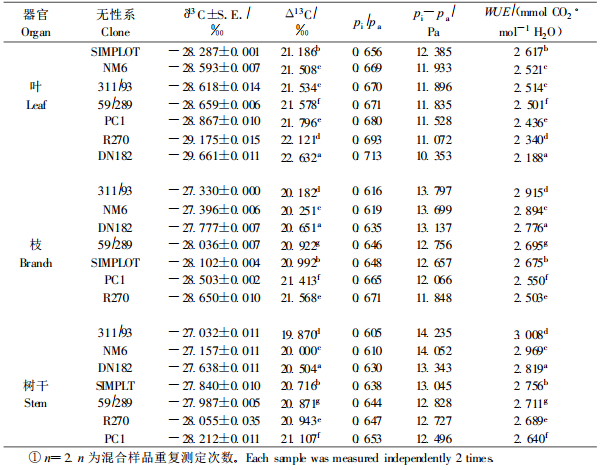
分别用叶、枝、干组织的Δ13C评价7个参试无性系的WUE,所得的结果不一致。用功能叶Δ13C得到的7个无性系的WUE由高到低的顺序是:SIMPLOT、NM6、311/93、59/289、PC1、R270、DN182;用当年生枝条Δ13C评价的顺序是:311/93、NM6、DN182、59/289、SIMPLOT、PC1、R270;而用树干Δ13C评价的顺序则是:311/93、NM6、DN182、SIMPLOT、59/289、R270、PC1(表 3)。可见用叶片Δ13C评价WUE的顺序与用气体交换法评价的结果基本一致,但与用枝Δ13C和干Δ13C评价的结果存在很大差异,尤其是SIMPLOT和DN182两个无性系。当年生枝和树干Δ13C求算的WUE的排列结果则基本一致。由于树干Δ13C反映了7个无性系从扦插到取样这一时间段的总水分利用效率平均值,因此,相对于叶和当年生枝条Δ13C得到的WUE而言,更具有代表性,更能反映7个无性系在大田条件下水分利用效率的高低。
不同无性系的pa-pi在11~1 4 Pa之间,pi/pa在0.6~0.7之间。pa-pi与气孔导度负相关,与光合系统活性正相关,由于没有不同时期气孔导度和光合系统活性的信息,无法判定不同无性系间pa-pi差异的具体原因。
2.4 不同无性系的Δ13C与生物量的关系由叶、枝、干的Δ13C与高、径生长量和生物量(表 4)的相关分析结果(表 5)可以看出:叶Δ13C与高、径生长量和生物量相关性不明显;枝Δ13C与1年生植株的树高、地径、干生物量、总生物量的相关性不显著,而与2年生植株的树高、胸径和2005年的树高生长量呈显著线性正相关。这是因为采样时间是2005年7月,枝样品为当年生枝,其Δ13C只能反映当年的生长情况。干Δ13C与1年生苗的树高、地径和生物量的相关性也不明显(表 5),但与2年生的树高达到极显著的正线性相关(表 5,图 2b),与当年树高生长量和胸径的相关性也达到显著水平(表 5,图 2a, c)。这说明枝、干Δ13C一方面能很好地反映杨树扦插苗在前2年的生长量,另一方面这种正相关性也说明在扦插苗的生长过程中,土壤水分是比较充足的。功能叶的Δ13C与2年的生长量均表现为不相关,其原因可能是由于叶片的形成时间短,其Δ13C只能反映这期间的生长情况和水分状况,而不能外推到整个生长季节和更长时间段。由于干Δ13C与树高间存在极显著的正线性相关(y=106.73x-1 546.2, R2=0.82, P<0.01),因此具有评价杨树在良好水分栽培条件下的速生性能的潜在价值。当然,这还需要在扩大研究样本容量的同时,对幼年植株(如1~3年生)的枝、干Δ13C与植株的高、径生长过程和生物量累积过程的关系开展进一步的研究。
|
|

|
|

|
图 2 杨树树干Δ13C与2年生植株生长量的相关性 Fig. 2 The relations between stem Δ13C and growth of 2-year-old poplar trees a. Δ13C与第2年树高生长量Δ13C versus height growth in the 2nd growth season; b. Δ13C与2年生树高Δ13C versus tree height at the end of the 2nd growth season;c. Δ13C与2年生胸径Δ13C versus tree diameter at breast height at the end of the 2nd growth season. |
用气体交换法测定植物的水分利用效率,适用于单叶水平的瞬时水分利用效率的测定,尤其适于研究引起水分利用效率快速变化的生理生态过程,如短时间内光照、叶温、气温、大气湿度、土壤含水量等变化对水分利用效率的影响及其生理机制。由于单叶的瞬时WUE在时间和空间上的变异很大且很复杂,受到诸多生态因子和植株生理状态的影响(万雪琴等, 2006),因此要将在单叶水平得到的瞬时水分利用效率外推到整株水平的长期水分利用效率是困难的(Bacon, 2004;张正斌, 2003),用其来直接评价作物或树木不同基因型的WUE高低存在一定的风险。植物组织干物质的稳定C同位素分辨率不仅可以用来实时评价植株的瞬时水分利用效率(Condon et al., 1990),更重要的是它提供了一种简单快捷的方法来测定植物的长期水分利用效率,而且还可以通过分析干标本和树木年轮(Bert et al., 1997; Leavitt et al., 2002)的δ13C来研究水分利用效率的历史变化。如果同时分析样品的氧同位素分辨率Δ18O,还可以进一步判定是气孔导度还是光合碳同化过程,或两者共同作用引起水分利用效率变化(Bacon, 2004; Farquhar et al., 1998; Price et al., 2002)。此法比气体交换法的优越之处是可以通过对长期积累于叶片或其他器官中的碳代谢产物的稳定碳同位素分析来评价叶片或植株生长过程中总的WUE特征,比用气体交换法测定的瞬时水分利用效率更有代表性(Hones, 2004; Bacon, 2004)。此外,此种测定不受时间和季节限制,样品采集烘干之后,其中的C同位素成分不再改变,故可以放置生长季后较空闲时测定。但是,目前昂贵的分析费用限制了它的应用潜力。
3.2 用气体交换法测定瞬时水分利用效率与C同位素分辨率测定长期水分利用效率结果不一致的原因本研究分别采用气体交换法和稳定碳同位素分辨率法测定了同一组杨树无性系的水分利用效率,所得结果很不一致。这说明用瞬时水分利用效率评价不同杨树基因型水分利用效率存在较大的局限性,其主要原因是瞬时水分利用效率在时间和空间尺度上的推绎困难。如本研究将典型晴天的瞬时水分利用效率日进程和中等光强的特定环境条件下不同叶片的瞬时WUE,外推到整株水平的长期水分利用效率,由于WUE同时受到基因型和环境条件的显著影响,本研究中特定条件下的瞬时WUE并不能完全代表实际多变环境条件下的长期WUE。因此,相对于瞬时水分利用效率来说,由于Δ13C覆盖时间和空间尺度更大,用其来评价不同基因型的长期水分利用效率更为可靠。
3.3 同一无性系不同时间段水分利用效率差异的原因本研究中所有7个无性系不同器官的Δ13C及相应的WUE存在明显差异。这种差异本质上反映了同一植株在生长过程中WUE在不同时间尺度上的变化。这种变化可以理解为在杨树生长过程中,多种生态因子综合影响气体交换过程的结果,而且在诸多的环境因子里,土壤有效水分含量起着突出作用(Bacon, 2004)。由于Δ13C与土壤有效水量正相关,与WUE负相关(Araus et al., 2003; Yu et al., 2005),因此推测本研究中同一无性系叶、枝、干Δ13C所反映出的不同时间尺度上的水分利用效率的差异的原因是:本研究中用于测定Δ13C的样品是雨季采集,其产生和生长过程中的土壤水分充足,空气湿度较大,因此其Δ13C较大,相应的WUE较小;在4—5月上旬和秋季的9—10月,由于降雨量较小,土壤和空气湿度较小,此期间生产的碳有机化合物的Δ13C较小,相应的WUE则较大,结果导致叶片的WUE低于枝,再低于干的现象。
3.4 Δ13C与产量间的关系Δ13C与产量间的关系较为复杂。以小麦为材料的大量研究表明,在水分条件充足时,Δ13C与生物产量呈正相关;在水分不足时,呈负相关;在中等水分条件或水分时而充足时而亏缺情况下,两者间的相关性不明显(Condon et al., 1987; Craufurd et al., 1991; Araus et al., 1999; Merah et al., 2002)。这是因为Δ13C同时受到气孔导度和光合活性的影响,并且在湿润的年份,作物产量受限于低的气孔导度,但在干旱的年份,低的气孔导度却成为一个有利因素。当其他因素(如叶片温度)被考虑时,情况会变得更加复杂,如当产量一定时,Δ13C与生长期正相关,与叶温负相关(Acevedo, 1993)。Araus等(2003)对水分输入量、平均温度、地表蒸发等环境因子对硬质小麦种实Δ13C和产量关系的影响的研究表明:土壤有效水分对Δ13C的影响最大;在产量低于2 000 kg·hm-2时,Δ13C与产量没有相关性;当产量从2 000 kg·hm-2增加到2 500 kg·hm-2时,Δ13C与产量的相关性急剧增强并在2 500 kg·hm-2时达到显著水平;当产量大于2 500 kg·hm-2时,产量与Δ13C间出现相对稳定的正相关关系。本研究的7个杨树无性系的枝、干组织的Δ13C与杨树的高径生长呈显著正相关。由于植物稳定C同位素组分的测定费用昂贵,本研究只分析了7个无性系的Δ 13C与生长量间的关系,所用样本量偏小。要更加准确地建立Δ13C与生长量间的关系,需要用更多的无性系样本对上述结论作进一步的验证;要确定苗期(如1年生或2年生)树干组织的Δ13C是否可作为杨树速生性的早期选择指标,还需要继续研究苗期组织的Δ13C与后期生长(如一个轮伐期结束时的生长量或产量)的关系,如果这种正相关性始终成立,则可将Δ13C作为良好水分栽培条件下速生杨树品种选育的早期选择指标。
万雪琴, 夏新莉, 尹伟伦, 等. 2006. 不同杨树无性系扦插苗水分利用效率的差异及其生理机制. 林业科学, 42(5): 133-137. |
张正斌. 2003. 作物抗旱节水的生理遗传育种基础. 北京: 科学出版社, 34-54.
|
赵凤君, 高荣孚, 沈应柏, 等. 2005. 水分胁迫下美洲黑杨不同无性系间叶片δ13C和水分利用效率的研究. 林业科学, 41(1): 36-41. |
Acevedo E. 1993.Potential of carbon isotope discrimination as a selection criteria in barley breeding//Ehleringer J R, et al. Stable isotopes and plant carbon-water relations. Academic Press, San Diego, CA: 399-417
|
Araus J L, Brown H R, Febrero A, et al. 1993. Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum. Plant, Cell and Environment, 16: 383-392. DOI:10.1111/j.1365-3040.1993.tb00884.x |
Araus J L, Amaro T, Zuhair Y, et al. 1997. Effect of leaf structure and water status on carbon isotope discriminationin field-grown durum wheat. Plant Cell and Environment, 20: 1484-1494. DOI:10.1046/j.1365-3040.1997.d01-43.x |
Araus J L, Slafer G A, Romagosa I. 1999. Durum wheat and barley yields in antiquity estimated from 13C discrimination of archaeological grains: a case study from the Western Mediterranean Basin. Australian Journal of Plant Physiology, 26: 345-352. |
Araus J L, Villegas D, Aparicio N, et al. 2003. Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions. Crop Science, 43(1): 170-180. DOI:10.2135/cropsci2003.1700 |
Bacon M A. 2004.Water use efficiency in plant biology//Bacon M A. Water use efficiency in plant biology. Blackwell CRC Press, Florida, USA, 1-26
|
Bert D, Leavitt S W, Dupouey J L. 1997. Variations of wood δ13C and water-use efficiency of Abies alba during the last century. Ecology, 78(5): 1588-1596. |
Condon A G, Richards R A, Farquhar G D. 1987. Carbon isotope discrimination is positively correlated with grain yield and dry matter production in field-grown wheat. Crop Science, 27: 996-1001. DOI:10.2135/cropsci1987.0011183X002700050035x |
Condon A G, Richards R A, Farquhar G D. 1992a. The effect of variation in soil water availability, vapour-pressure deficit and nitrogen nutrition on carbon isotope discrimination in wheat. Australian Journal of Agricultural Research, 43: 935-947. DOI:10.1071/AR9920935 |
Condon A G, Richards R A. 1992b. Broad sense heritability and genotype environment interaction for carbon isotope discrimination in field grown wheat. Australian Journal of Agricultural Research, 43: 921-934. |
Condon A G, Farquhar G D, Richards R A. 1990. Genotypic variation in carbon-isotope discrimination and transpiration efficiency in wheat: leaf gas exchange and whole plant studies. Australian Journal of Plant Physiology, 17: 9-22. |
Craufurd P Q, Austin R B, Acevedo E, et al. 1991. Carbon isotope discrimination and grain-yield in barley. Field Crops Research, 27: 301-314. DOI:10.1016/0378-4290(91)90038-W |
Farquhar G D, Richards R A. 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Australian Journal of Plant Physiology, 11: 539-553. |
Farquhar G D, Barhour M M, Henry B K. 1998.Interpretation of oxygen isotope composition of leaf material//Griffiths H. Stable isotope: integrated of biological, ecological and geochemical process. BIOS Scientific Publishers, Oxford, 27-62
|
Farquhar G D, Ehleringer J R, Hubrick K T. 1989. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
Hall A E, Richards R A, Condon A G, et al. 1994.Carbon isotope discrimination and plant breeding//Janick J. Plant breeding review: Vol 12.John Wiley & Son, New York, 81-113
|
Hones H. 2004.What is water use efficiency//Bacon M A. Water use efficiency in plant biology. Blackwell CRC Press, Florida, USA, 27-40
|
Hubick K T, Farquhar G D. 1989. Carbon isotope discrimination and the ratio of carbon gained to water lost in cultivars of barley. Plant, Cell and Environment, 12: 795-804. DOI:10.1111/j.1365-3040.1989.tb01641.x |
Impa S M, Nadaradjan S, Boominathan P, et al. 2005. Carbon isotope discrimination accurately reflects variability in WUE measured at a whole plant level in rice. Crop Science, 45(6): 2517-2522. DOI:10.2135/cropsci2005.0119 |
Ismail A M, Hall A E. 1992. Inheritance of carbon isotope discrimination and water use efficiency in cowpea. Crop Science, 33: 498-503. |
Lambrides C J, Chapman S C, Shorter R. 2004. Genetic variation for carbon isotope discrimination in sunflower: association with transpiration efficiency and evidence for cytoplasmic inheritance. Crop Science, 44(5): 1642-1653. DOI:10.2135/cropsci2004.1642 |
Leavitt S W, Wright W E, Long A. 2002. Spatial expression of ENSO, drought, and summer monsoon in seasonal δ13C of ponderosa pine tree rings in southern Arizona and New Mexico. Journal of Geophysical Research-Atmospheres, 107: 43-49. |
Marshall J D, Zhang J W. 1994. Carbon isotope discrimination and water-use efficiency in native plants of the north-central Rockies. Ecology, 75(7): 1887-1895. DOI:10.2307/1941593 |
Matus A, Slinkard A E, van Kassel C. 1996. Carbon isotope discrimination and indirect selection for transpiration efficiency at flowering in lentil (Lens culinaris Medikus), spring wheat (Triticum aestivum L.), durum wheat (T. turgidum L.) and canola (Brassica napus L.). Euphytica, 87: 141-151. |
Merah O, Delens E, Souyris I, et al. 2002. Stability of carbon isotope discrimination and grain yield in durum wheat. Crop Science, 41: 677-681. |
Price A H, Cairns J E, Horton P, et al. 2002. Linking drought resistance mechanisms to drought avoidance in upland rice using QTL approach: progress and new opportunities to integrate stomatal and mesophyll responses. Journal of Experimental Botany, 53: 989-1004. DOI:10.1093/jexbot/53.371.989 |
Read J J, Johnson D A, Asay K H, et al. 1991. Carbon isotope discrimination, gas exchange and WUE in crested wheat grass clones. Crop Science, 31: 1203-1208. DOI:10.2135/cropsci1991.0011183X003100050025x |
Rebetzke G J, Condon A G, Richards R A, et al. 2002. Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Science, 42: 739-745. |
Richards R A, Condon A G. 2002.Challenges ahead in using carbon isotope discrimination in plant breeding program//Ehleringer J R, et al. Stable isotopes and plant carbon-water relations. Academic Press, New York, 451-462
|
Richards R A. 1996. Defining selection criteria to improve yield under drought. Plant Growth Regulation, 20: 157-166. DOI:10.1007/BF00024012 |
Sheshshayee M S, Bindumadhava H, Shankar A G, et al. 2003. Breeding strategies to exploit water use efficiency for crop improvement. Journal of Plant Biology, 30(2): 253-268. |
Stiller W N, Read J J, Constable G A, et al. 2005. Selection for water use efficiency traits in a cotton breeding program: cultivars differences. Crop Science, 45(3): 1107-1113. DOI:10.2135/cropsci2004.0545 |
Udayakumar M, Sheshshayee M S, Nataraj K N, et al. 1998. Why breeding for water use efficiency has not been successful: An analysis and alternate approach to exploit this trait for crop improvement. Current Science, 74(11): 994-1000. |
Yu Mei, Xie Yichun, Zhang Xinshi. 2005. Quantification of water use efficiency along a moisture gradient in Northeastern China. Journal of Environmental Quality, 34(4): 1311-1318. |