 2007, Vol. 43
2007, Vol. 43文章信息
- 吕瑜良, 刘世荣, 孙鹏森, 张国斌, 张瑞蒲.
- Lü Yuliang, Liu Shirong, Sun Pengsen, Zhang Guobin, Zhang Ruipu.
- 川西亚高山暗针叶林叶面积指数的季节动态与空间变异特征
- Seasonal and Spatial Variations of Leaf Area Index of Sub-Alpine Dark Coniferous Forest during Growing Season in Western Sichuan
- 林业科学, 2007, 43(8): 1-7.
- Scientia Silvae Sinicae, 2007, 43(8): 1-7.
-
文章历史
- 收稿日期:2006-10-20
-
作者相关文章



2. 西南大学地理科学学院 重庆 400715;
3. 山东省烟台市昆嵛山林场 烟台 264000
2. School of Geography Science, Southwest University Chongqing 400715;
3. Kunyushan Forest Field, Yantai, Shandong Province Yantai 264000
叶面积指数(LAI)的定义有多种(Asner et al., 2003),最近文献中使用最多的为“单位面积上叶片的倾斜投影面积”(Barclay, 1998)。Chen等(1992)将针叶林LAI定义为“单位面积上总针叶投影面积的一半”,即投影系数为0.5。LAI能够定量描述植被冠层结构及其动态变化,反映冠层结构诸多影响因素的综合作用(Chen et al., 1997),是植被系统的重要属性之一(Lee et al., 2004),控制着植被生态系统中许多物理和生物地球化学过程(如初级生产、蒸腾作用、能量交换、碳与养分循环以及降雨截留等)(Asner et al., 2003)。因此许多领域都将LAI作为一个研究植被相关过程的关键变量使用(Asner et al., 2003; 王希群等, 2005),许多气候模型和生态模型也把它作为主要的输入项(Running et al., 1988; Boman, 1993; Sellers et al., 1997)。
川西(包括四川省的凉山、阿坝和甘孜3个自治州)位于长江上游,地处青藏高原东南缘和西北干旱草原区向东南湿润森林区的过渡地带,地形复杂,山高坡陡,河谷深切,地震活跃,岩体松散,极易发生崩塌,泥石流灾害频繁(李承彪, 1990),是一个地理位置特殊、自然环境复杂、生态系统脆弱的典型区域(姚建等, 2004)。亚高山暗针叶林是该区的主要植被类型,多沿河曲与谷坡呈树枝状分布,在涵养水源、保持水土、调节气候、维护生态平衡等方面起着巨大的作用(李承彪, 1990; 刘庆, 2002; 管中天, 2005)。目前,关于川西亚高山暗针叶林LAI的研究有些报道,如Luo等(1997)测定了贡嘎山不同海拔高度云冷杉林的最大LAI,Lü等(2002)研究了青藏高原东南部植被LAI与初级生产力之间的关系,李崇巍等(2005)利用LAI与归一化植被指数(NDVI)之间的关系,反演了岷江上游主要植被类型的LAI及其空间分布格局。这些研究提供了该区宏观尺度上主要植被类型的LAI,但对该区主要森林类型的亚高山暗针叶林LAI的季节变化、空间分异和群落类型变化的规律及机制的定量研究很少。本文试图应用Li-cor LAI-2000植物冠层分析仪,通过大量的实测分析,研究亚高山暗针叶林LAI时空变化特征及其影响因素,以揭示LAI季节变化规律及其群落类型之间的差异性、LAI随海拔和坡向变化的空间格局及LAI在林分尺度和小流域尺度上的变异性,为确定川西亚高山暗针叶林影响水文循环过程的植被结构特征参数,建立植被生态过程与水文过程的耦合关系,以及预测森林植被对全球变化的响应提供依据。
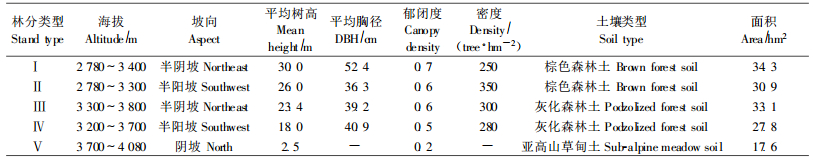
1 研究区概况研究区位于四川卧龙亚高山暗针叶林生态系统定位研究站(30°51′41″ N,102°58′21″ E)。试验小流域距离该站约300 m,面积约1.44 km2,海拔2 780~4 080 m。属青藏高原气候区,典型的亚热带内陆山地气候。12月份气温最低,平均为-5.2 ℃,7月份最高,平均为12.4 ℃,年均气温4.3 ℃。夏季相对湿度较大,为84.8%,冬季较小,为75.5%,年均湿度79%。年降水848.9 mm,集中于6—9月份,占全年降水的64.25%。风向以北风及东北风为主。年蒸发量772.5 mm,日照时数1 185.4 h。小流域的植被主要为岷江冷杉(Abies faxoniana)原始林,包括以下4个林分类型:藓类-箭竹-岷江冷杉林(moss-Fargesia nitida-Bashania fangiana-A. faxoniana stand)、草类-箭竹-岷江冷杉林(grass-F. nitida-B. fangiana-A. faxoniana stand)、藓类-杜鹃-岷江冷杉林(moss-Rhododendron spp.-A. faxoniana stand)和草类-杜鹃-岷江冷杉林(grass-Rhododendron spp.-A. faxoniana stand),代号依次为Ⅰ、Ⅱ、Ⅲ、Ⅳ;其次为杜鹃(Rhododendron spp.)灌丛,代号为V(表 1)。
|
|
LAI用植物冠层分析仪(Li-cor LAI-2000 Plant Canopy Analyzer)测定。样点布设主要考虑植被和地形2个因子。植被包括小流域的全部4个岷江冷杉林分和1个杜鹃灌丛,海拔范围为2 800 ~3 900 m,坡向分半阴坡和半阳坡。从小流域沟口到顶端,沿两边坡面,结合群落类型,共26个样点,其中藓类-箭竹-岷江冷杉林7个,草类-箭竹-岷江冷杉林6个,藓类-杜鹃-岷江冷杉林和草类-杜鹃-岷江冷杉林各5个,杜鹃灌丛3个。每个样点观测3组数据,每组数据测定10个LAI值,取其平均值,每次共测得78组数据。观测高度距地面1 m。观测时间为2005年7—9月份,每月观测2~3次,每次观测的样点位置相同。
2.2 数据处理与分析杜鹃灌丛LAI为冠层分析仪实测值,冷杉林LAI按照下列公式(Chen, 1996)计算:
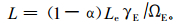
|
式中:L为实际叶面积指数;α为树干等非树叶因素占总面积的比率;Le为有效叶面积指数,可以由LAI-2000直接测定;γE为不同针叶树种的针叶总面积与簇面积的比率,对于阔叶树种,γE取值为1,对针叶树种,γE取值为1.4 (Chen, 1996);ΩЕ是针叶聚集指数,利用跟踪辐射与冠层结构测量仪(TRAC)对4种冷杉林分类型的测定值约为0.95,并且在不同林分类型之间没有显著性差异。
测得的各群落类型LAI为该类型该次所有观测点的算术平均值,小流域LAI为该5种群落类型LAI的面积加权平均值。把测得的所有LAI数据按照观测时间、群落类型、海拔、坡向进行分类统计,对各类数据进行单因素方差分析,并对4个岷江冷杉林分的LAI两两之间进行相关分析及均值比较。数值计算用EXCEL软件,统计分析用SPSS软件。
3 结果与分析 3.1 生长季节小流域平均LAI的变化规律研究期间,小流域LAI随时间的变化为单峰曲线,峰值出现在8月中旬,为5.82±1.32,7月初至8月中旬LAI约增加25%,8月中旬至9月末约减小25%。方差分析结果表明,7—9月LAI的差异极显著(p < 0.001)(图 1)。说明研究期间,小流域LAI存在明显的时间动态变化。

|
图 1 生长季节小流域LAI的时间动态 Fig. 1 Temporal dynamic of LAI in the small catchment in growing season |
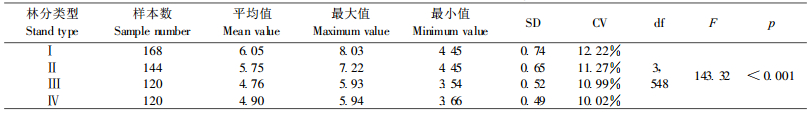
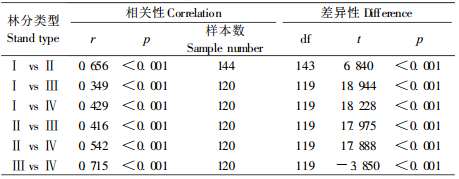
小流域4种岷江冷杉林分LAI的平均值、标准差和变异系数都是箭竹-岷江冷杉林较大,杜鹃-岷江冷杉林较小,其中藓类-箭竹-岷江冷杉林最大,相应为6.05,0.7385和12.22%。方差分析结果表明,不同林分之间LAI的差异极显著(p < 0.001)(表 2)。均值比较表明,4种岷江冷杉林分LAI两两之间差异都极显著(p < 0.001),其中箭竹-岷江冷杉林与杜鹃-岷江冷杉林之间的差异性较大,相关性较弱,2种杜鹃-岷江冷杉林之间和2种箭竹-岷江冷杉林之间的差异性较小,相关性较强(表 3)。说明分布在小流域不同海拔、不同坡向的岷江冷杉林间LAI差异较大,灌木层优势种不同是造成差异的主要原因。
|
|
|
|
4种岷江冷杉林分LAI的时间变化趋势一致,但变化幅度不同。从7月初到8月中旬,LAI的增加幅度为箭竹-岷江冷杉林较大,杜鹃-岷江冷杉林较小,其中藓类-箭竹-岷江冷杉林最大,草类-杜鹃-岷江冷杉林最小;从8月中旬到9月末,LAI的减小幅度为藓类-杜鹃-岷江冷杉林最大,草类-杜鹃-岷江冷杉林最小(图 2)。

|
图 2 生长季节不同岷江冷杉林分LAI的时间动态 Fig. 2 Temporal dynamics of LAI for different A. faxoniana stands in growing season |
生长季节,随着海拔升高,小流域LAI先增大,后减小,转折点出现在3 000 m左右,LAI为6.17±0.53,减幅随海拔升高越来越大,3 900 m处LAI最小,为2.69±0.17。LAI(y)与海拔(x)之间的回归方程为: y =-22.408 6+0.019 6x-0.000 003 360 1x2 (R2 = 0.999 6, p < 0.001, n=12)。方差分析结果表明,不同海拔之间LAI的差异极显著(p < 0.001)(图 3)。说明海拔是LAI的一个重要影响因子。

|
图 3 生长季节小流域LAI随海拔升高的变化规律 Fig. 3 Changes in LAI with ascending altitude in the catchment in growing season |
不同海拔LAI的时间动态变化趋势基本一致,与小流域LAI的时间动态相同。但是,低海拔地区的曲线峰度比高海拔大(图 4)。

|
图 4 生长季节小流域不同海拔高度LAI的时间动态 Fig. 4 Temporal dynamic of LAI at different altitudes in the small catchment in growing season |
不同海拔LAI的变异系数为7.64%~23.09%。随着海拔升高,LAI的变异性变化比较复杂。总体上,LAI的变异性在低海拔较小,在高海拔较大。其中,海拔3 500 m最小,为7.64%,海拔3 800 m最大,为23.09%(图 5)。

|
图 5 生长季节小流域不同海拔高度LAI的变异系数 Fig. 5 Coefficient of variability (CV) of LAI at different altitudes in the small catchment in growing season |
研究期间,小流域半阴坡LAI平均值(5.51)比半阳坡(5.36)稍大。方差分析结果表明,两者之间差异显著(p < 0.05)(表 4)。说明坡向也是LAI的一个主要影响因子。
|
|

不同坡向LAI的时间变化趋势大致相同,与小流域的时间动态一致,但在时间上存在差异。7月初,LAI半阴坡比半阳坡低,到7月中旬,前者逐渐超过后者,8月中旬两者之间相差最大,此后,两者虽然都逐渐减小,但直到9月底,前者始终比后者大(图 6)。

|
图 6 生长季节小流域不同坡向LAI的时间动态 Fig. 6 Temporal dynamic of LAI at different aspects in the small catchment in growing season |
虽然针叶林是常绿的,但其LAI也存在季节变化(Chen, 1996)。从卫星遥感影像上可以看到,针叶林的植被指数在夏季最大,冬季最小(Cihlar, 1996)。Chen(1996)认为,针叶林LAI的季节变化在25%~30%之间,依赖于针叶的平均寿命以及当年和前年针叶的数量。Luo等(2002)研究表明,LAI的季节变化主要受控于平均温度、降水量和辐射的季节变化。本研究结果表明,生长季节川西亚高山暗针叶林LAI的时间动态变化比较明显,为单峰曲线,峰值在8月中旬。这与我国长白山温带山地云冷杉(Picea spp.和Abies spp.)林、针阔混交林(Luo et al., 2002;周宇宇等,2003)以及加拿大北部老龄黑杉(Picea mariana)林(75~90年生) (Chen, 1996)的变化趋势基本一致。研究期间,LAI增加(从7月初到8月中旬)与减小(从8月中旬到9月末)的幅度相当,约为25%。这与Chen(1996)估计的年变化接近。需要指出的是,本研究中LAI的观测高度为1.0 m,25%的变化包括乔木层和灌木层的变化。Wirth等(2001)对热带雾林的研究表明,LAI存在明显的垂直分布,下层林冠的LAI较小,5 m以上林冠LAI占总LAI的50%以上。亚高山暗针叶林冠层结构虽然没有热带林的复杂,但岷江冷杉林原始林为复层异龄林,林下灌木、幼树生长良好,对LAI的贡献较大。对于川西亚高山暗针叶林LAI的垂直分布和全年变化规律,今后还需要做进一步的观测研究。
4.2 不同岷江冷杉林分之间LAI的差异性植被LAI主要受物种组成、结构、立地条件、微气象等许多因素的影响(Asner et al., 2003; Jonckheere et al., 2004),也与叶片的数量、分布角度及空间分布有关。箭竹-岷江冷杉林乔木层的平均高度、平均胸径、林分郁闭度都比杜鹃-岷江冷杉林大,灌木层优势种前者为冷箭竹,后者为杜鹃,冷箭竹密度和盖度比杜鹃的大,前者林内比后者阴暗,故前者LAI比后者的大。分布在不同海拔、不同坡向的岷江冷杉林,由于林分的组成、结构、生长状况及微气象条件等不同,导致LAI差异极为显著。箭竹-岷江冷杉林与杜鹃-岷江冷杉林之间的差异主要是由灌木层的组成不同引起的,箭竹-岷江冷杉林之间、杜鹃-岷江冷杉林之间的差异主要是由不同坡向微气象条件不同引起的。相对而言,由林分组成和结构不同引起的LAI的差异性比由微气象条件不同引起的差异性要大。
4.3 高山峡谷区地形对LAI的影响关于LAI与海拔之间关系的研究报道并不多。Luo等(2002)在四川贡嘎山测定了不同海拔森林的最大LAI,但没有揭示出LAI与海拔之间的关系。李崇巍等(2005)分析了岷江上游植被指数(NDVI)与海拔之间的关系,海拔范围为1 500~3 200 m,NDVI随着海拔升高而增加,由LAI与NDVI正相关(R2=0.807 1),可推断LAI随着海拔升高而增加,但对更高海拔范围没有进行研究。本研究结果表明,海拔是影响LAI的一个重要因子。在海拔3 000 m以下,LAI随海拔的升高而增加,与李崇巍等(2005)的研究结果一致。在海拔3 000 m以上,LAI随海拔升高而减小,且幅度越来越大。研究表明,在四川卧龙地区,海拔每升高100 m,降雨量增加3.4%左右,蒸发量下降2.8%,气温下降0.6 ℃,相对湿度增加2.4%。随着海拔升高,年均温度下降,年降水量增加,水热条件逐渐好转,植被生长越来越好;但随着海拔继续升高,水热条件逐渐变差,造成植被生长变差(李景文, 1994)。小流域下部为生长茂密的箭竹-岷江冷杉林,中部为生长较好的杜鹃-岷江冷杉林,上部为生长稀疏的杜鹃灌丛,符合这一规律,可以解释LAI随海拔升高的变化规律。但转折点在海拔3 000 m处,是否在这个高度试验小流域的暗针叶林生长状况最好,调查资料没有显示出来,已有的文献也没有说明川西亚高山暗针叶林在海拔3 000 m处生长最好。因此,这个数据是否适合整个川西亚高山暗针叶林,尚须进一步的研究证实。
本研究表明,坡向也是影响LAI的主要地形因子。这主要是因为不同坡向的小气候差异很大,导致不同坡向上植被类型及其生长状况明显不同(卧龙自然保护区管理局,1987)。半阴坡降水量、相对湿度比半阳坡大,前者日照时数、太阳辐射、平均温度和蒸发量比后者小。岷江冷杉喜阴耐湿,在半阴坡生长比半阳坡好(李承彪,1990;刘庆,2002;管中天,2005),林分平均树高、郁闭度半阴坡都比半阳坡大(表 1),因而前者LAI比后者大。生长季初期,半阳坡林内光照比半阴坡充足,温度高,林下植物生长快,LAI比半阴坡大,但进入生长季中后期,半阴坡植物生长逐渐赶上并超过半阳坡,LAI比半阳坡大。
除海拔和坡向外,坡度也是一个重要的地形因子。但本文没有分析其与LAI之间的关系,是否不同坡度LAI存在明显的变化规律,还有待研究。
4.4 亚高山暗针叶林小流域LAI的变异性本研究表明,观测期间,川西岷江冷杉林LAI的变异系数在林分尺度上为10.02%~12.22%,在小流域尺度上为19.79%~22.80%,前者约是后者的1/2。LAI主要受植被、地形、气候等因子的影响(Asner et al., 2003; Jonckheere et al., 2004)。在林分尺度上,上述因子基本上相同,故LAI的变异性较小,但在流域尺度上,则存在一定的异质性,故LAI的变异性较大。Asner等(2003)的研究表明,全球跨越干旱、半干旱、温带、热带、寒带以及人工生态系统的15个生物区系(biomes)LAI变化范围很大,温带和热带常绿阔叶林变异系数最小,为2%~20%,草地、灌丛及苔原最大,为70%~78%,寒温性常绿针叶林为30%~40%。川西岷江冷杉林LAI的变异性在林分尺度上约是全球寒温性常绿针叶林的1/3。据此可以推断,随着空间尺度的增大,亚高山暗针叶林LAI的变异性增大。Aragao等(2005)在亚马逊东部热带雨林的研究表明,LAI的变异系数在0.25 hm2上为5.2%~23%,在0.75 hm2上为1.8%~12%,随着空间尺度增大,LAI的变异系数减小。这个结论似乎与本研究相反,但并不矛盾。本研究的结论是通过比较暗针叶林在林分(15~35 hm2)尺度、小流域(1.44 km2)尺度和全球尺度上LAI的变异性大小得出的,而Aragao等(2005)的结论是通过比较0.25和0.75 hm2尺度上LAI的变异性大小得出的。不难理解,热带森林植物种类非常丰富,面积越小,植被的异质性越大,LAI的变异性也越大。而在亚高山暗针叶林和寒温性针叶林,植物种类组成及结构相对较为简单,在较小的空间尺度上,植被状况基本相同,在较大的空间尺度上,植被才会表现出变异性。本文没有研究亚高山暗针叶林1 hm2以下LAI的异质性随空间尺度的变化,也未找到在大尺度上热带雨林LAI的变异性的数据,因而无法更好地比较亚高山暗针叶林LAI的变异性与热带雨林的异同。
亚高山暗针叶林LAI的变异系数随着海拔升高变化比较复杂。低海拔处变异系数较小,高海拔处变异系数较大,海拔3 700 ~3 800 m变异系数最大。不同坡向之间,变异系数半阴坡比半阳坡大,这主要与不同坡向之间微气象条件的差异有关。比较不同海拔与不同坡向的变异系数,在低海拔,不同坡向之间的变异系数比不同海拔之间大,但在高海拔,则相反。因此,在低海拔测定暗针叶林LAI时,坡向比海拔的影响大,在高海拔则相反。
管中天. 2005. 森林生态研究与应用. 成都: 四川科技出版社.
|
李承彪. 1990. 四川森林生态研究. 成都: 四川科技出版社.
|
李崇巍, 刘世荣, 孙鹏森, 等. 2005. 岷江上游植被冠层降水截留的空间模拟. 植物生态学报, 29(1): 60-67. DOI:10.3321/j.issn:1005-264X.2005.01.009 |
李景文. 1994. 森林生态学. 北京: 中国林业出版社.
|
刘庆. 2002. 亚高山针叶林生态学研究. 成都: 四川大学出版社.
|
王希群, 马履一, 贾忠奎, 等. 2005. 叶面积指数的研究和应用进展. 生态学杂志, 24(5): 537-541. DOI:10.3321/j.issn:1000-4890.2005.05.015 |
卧龙自然保护区管理局. 1987. 卧龙植被及资源植物. 成都: 四川科学出版社.
|
姚建, 丁晶, 艾南山. 2004. 岷江上游生态脆弱性评价. 长江流域资源与环境, 13(4): 380-383. DOI:10.3969/j.issn.1004-8227.2004.04.016 |
周宇宇, 唐世浩, 朱启疆, 等. 2003. 长白山自然保护区叶面积指数测量及其结果. 资源科学, 25(5): 38-42. |
Aragao L E O S, Shimabukuro Y E, Santo F D B E, et al. 2005. Landscape pattern and spatial variability of leaf area index in Eastern Amazonia. For Ecology and Management, 211: 240-256. DOI:10.1016/j.foreco.2005.02.062 |
Asner G P, Scurlock J M O, Hicke J A. 2003. Global sythesis of leaf area index observation: Implications for ecological and remote sensing studies. Globe Ecology and Biogeography, 12: 191-205. DOI:10.1046/j.1466-822X.2003.00026.x |
Barclay H G. 1998. Conversion of total leaf area to projected leaf area in lodgepole pine and Douglas-fir. Tree Physiology, 18: 185-193. DOI:10.1093/treephys/18.3.185 |
Boman G B. 1993. Importance of leaf area index and forest type when estimating photosynthesis in boreal forests. Remote Sensing Environment, 43: 303-314. DOI:10.1016/0034-4257(93)90072-6 |
Chen J M. 1996. Optically-based methods for measuring seasonal variation in leaf area index of boreal conifer forests. Agriculture and Forest Meteorology, 80: 135-136. DOI:10.1016/0168-1923(95)02291-0 |
Chen J M, Black T A. 1992. Defining leaf-area index for non-flat leaves. Plant Cell Environment, 15: 421-429. DOI:10.1111/pce.1992.15.issue-4 |
Chen J M, Rich P M, Gower S T, et al. 1997. Leaf area index of boreal forests: Theory, techniques, and measurements. Journal of Geophysical Research, 102: 29429-29443. DOI:10.1029/97JD01107 |
Cihlar J. 1996. Identification of contaminated pixels in AVHRR composite images for studies of land biosphere. Remote Sensing Environment, 55: 153-162. DOI:10.1016/0034-4257(95)00195-6 |
Jonckheere I, Fleck S, Nackaerts K, et al. 2004. Review of methods for in situ leaf area index determination Part Ⅰ. Theories, sensors and hemispherical photography. Agriculture and Forest Meteorology, 121: 19-35. DOI:10.1016/j.agrformet.2003.08.027 |
Lü J H, Ji J J. 2002. A simulation study of atmosphere-vegetation interaction over the Tibetan Plateau. part Ⅰ: Net primary productivity and leaf area index. Chinese Journal of Atmosphere Science, 26: 255-262. |
Lee K S, Cohen W B, Kennedy R E, et al. 2004. Hyperspectral versus multispectral data for estimating leaf area index in four different biomes. Remote Sensing Environment, 91: 508-520. DOI:10.1016/j.rse.2004.04.010 |
Luo T X, Li W H, Zhao S D. 1997. Distribution patterns of leaf area index for major coniferous forest types in China. Journal of Chinese Geography, 7: 61-73. |
Luo T X, Neilson R P, Tian H Q, et al. 2002. A model for seasonality and distribution of leaf area index of forests and its application to China. Journal of Vegetation Science, 13: 817-830. DOI:10.1111/jvs.2002.13.issue-6 |
Running S W, Coughlan J C. 1988. A general model of forest ecosystem processes for regional applications. Ⅰ. Hydrologic balance, canopy gas exchange and primary production processes. Ecological Modelling, 42: 125-154. DOI:10.1016/0304-3800(88)90112-3 |
Sellers P J, Dickinson R E, Randall D A, et al. 1997. Modeling the exchanges of energy, water and carbon between continents and the atmosphere. Science, 275: 502-509. DOI:10.1126/science.275.5299.502 |
Wirth R, Weber B, Ryel R J. 2001. Spatial and temporal variability of canopy structure in a tropic moist forest. Acta Oecologica, 22: 1-10. DOI:10.1016/S1146-609X(00)01093-6 |