 2007, Vol. 43
2007, Vol. 43文章信息
- 马姜明, 刘世荣, 史作民, 张远东, 康冰, 陈宝玉.
- Ma Jiangming, Liu Shirong, Shi Zuomin, Zhang Yuandong, Kang Bing, Chen Baoyu.
- 川西亚高山暗针叶林恢复过程中群落物种组成和多样性的变化
- Changes of Species Composition and Diversity in the Restoration Process of Sub-Alpine Dark Brown Coniferous Forests in Western Sichuan, China
- 林业科学, 2007, 43(5): 17-23.
- Scientia Silvae Sinicae, 2007, 43(5): 17-23.
-
文章历史
- 收稿日期:2006-06-28
-
作者相关文章


2. 西北农林科技大学生命科学学院 杨凌 712100;
3. 北京师范大学生命科学学院 北京 100875
2. College of Life Sciences, Northwest Sci-Tech University of Agriculture and Forestry Yangling 712100;
3. College of Life Sciences, Beijing Normal University Beijing 100875
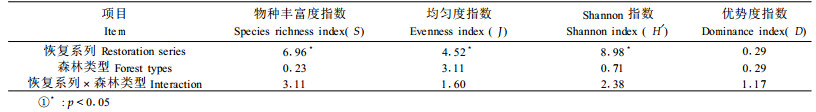
群落多样性是群落生态学研究乃至整个生态学研究中十分重要的内容(史作民等,2002)。任何群落或生态系统类型都有物种多样性特征,而这种特征是该生态系统功能维持的生物基础(Tilman et al., 1994; Grime, 1997; 张继义等,2004)。对物种多样性的研究可体现群落和生态系统的结构类型、组织水平、发展阶段、稳定程度、生境差异等(阎海平等, 2001)。物种多样性恢复成为生态系统恢复过程的重要特征(林开敏等, 2001)。维持和发展生物多样性成了生态系统恢复的一个主要目标(Young, 2000; Zerbe et al., 2006)。在许多恢复实践中也把多样性作为评价恢复成功的主要指标之一(Ruiz-Jaen et al., 2005)。对于物种组成和多样性的研究主要从海拔梯度、森林恢复过程与物种多样性的关系等方面进行(贺金生等, 1997; Guariguata et al., 1997; 温远光, 1998; Vázquez et al., 1998; 吴彦等, 2001; 包维楷等, 2002; 赵常明等, 2002; 郝占庆等, 2002a; 石胜友等)。
川西亚高山林区地处青藏高原东南缘,是世界生物多样性研究的热点地区(杨万勤等, 2003),亚高山暗针叶林是该区原有的主要森林类型(张远东等, 2005a)。自20世纪50年代中期以来,该林区的天然暗针叶林被大规模采伐,导致植被覆盖率急剧减小,部分物种消失,生态系统退化以及生态功能减弱(刘庆等, 2001)。亚高山暗针叶林植被恢复成为一项十分必要和紧迫的任务。近年来该区大部分研究主要集中在人工林结构与功能的变化,而对暗针叶林采伐后形成的不同恢复系列次生林的结构变化研究较少,仅涉及林地水文效应方面(张远东等, 2005b)。研究川西亚高山暗针叶林不同自然恢复系列群落物种组成和多样性的变化,对于揭示其生态学过程与机制,正确评价森林恢复的进程以及加快退化暗针叶林生态系统恢复具有十分重要的意义。本文主要应用α, β多样性指数分析了川西亚高山暗针叶林不同恢复系列物种多样性特征及其变化规律,以便为川西亚高山地区退化天然暗针叶林的生物多样性保育和生态系统综合管理提供参考。
1 研究区概况研究区位于四川省理县米亚罗林区(31°24′—31°55′ N,102°35′—103°4′ E), 属于青藏高原向四川盆地过渡的高山峡谷区,海拔2 200~5 500 m,典型的高山峡谷地貌。该区夏季温凉多雨,冬季寒冷干燥,以海拔2 760 m的米亚罗镇为例,全年降水量700 ~1 000 mm,年蒸发量1 000~1 900 mm,1月均温-8 ℃,7月均温12.6 ℃,≥10 ℃年积温为1 200~1 400 ℃,属季风性山地气候。
米亚罗林区植被垂直成带明显,其类型和生境随海拔及坡向而分异(蒋有绪, 1963a)。海拔2 700 m以下为亚高山针阔混交林带;2 700~4 000 m为亚高山针叶林及高山疏林带;4 000 m以上为高山草甸、高山荒漠及积雪带(蒋有绪, 1963b)。亚高山暗针叶林带中,在海拔3 100~3 500 m的阴坡、半阴坡上分布着箭竹(Sinarundinaria chungii)针叶林环的各种林型;在海拔3 300~3 600 m的阴坡、半阴坡,以及河谷底部的潮湿地方分布着藓类针叶林环的各种林型。这2个林环组成亚高山暗针叶林带内的主要森林类型。大多数情况下,同一林型组,甚至同一林环的不同林型,受人为破坏后,迹地上形成的植被在较长时间内其演替过程大致是相似的。箭竹针叶林环和藓类针叶林环遭受破坏(如采伐)后分别形成箭竹红桦(Betula albo-sin ensis)和藓类红桦林1)。
1) 中国林业科学研究院.1963.西南高山村区森林综合考察报告
米亚罗林区在1950—1978年间进行过大规模采伐, 之后采伐量逐年减少, 至1998年停采封育,主要伐区位于海拔2 800~3 600 m处(张远东等,2005b),构成了不同恢复系列的次生林群落类型,所形成的次生林主要分布在阴坡、半阴坡, 采伐前是亚高山原始冷杉林的集中分布区(张远东等,2005a)。这为研究退化暗针叶林的自然恢复提供了理想的演替系列。原始暗针叶老龄林主要优势种为岷江冷杉(Abies faxoniana),次生林优势种主要为红桦。主要灌木有箭竹、杜鹃(Rhodoendron spp.)、蔷薇(Rosa spp.)、卫矛(Euonymus spp.)、忍冬(Lonicera spp.)、茶藨子(Ribes spp.)、栒子(Cotoneaster spp.)、小檗(Berberis spp.)、五加(Acanthopanax gracilithlus)等。主要草本植物有鹿药(Smilacina japonica)、扭柄花(Streptopus obtusatus)、鹿耳韭(Allium ovalifolium)、蝎子草(Girardinia suborbiculata)、菝葜(Smilax spp.)、细辛(Asarum himalaicum)、高山露珠草(Circaea alpina)、蟹甲草(Parasenecio spp.)、唐松草(Thalictrum spp.)、草玉梅(Anemone rivularis)、山酢酱草(Oxalis griffitii)、七筋姑(Clintonia udensis)等。米亚罗森林土壤主要为山地棕壤。
2 研究方法 2.1 样地调查分别选择20世纪50、60、70、80年代暗针叶林采伐后恢复的天然次生林为对象,采用以空间代替时间的方法,在海拔3 100~3 600 m的阴坡、半阴坡,立地条件基本一致的生境,分别对箭竹红桦林(海拔3 100~3 500 m)和藓类红桦林(海拔3 300~3 600 m) 2种森林类型进行群落调查,每种类型设置20、30、40和50年生的恢复系列,采用典型取样法设置50个标准地,2种林型中,20、30、40和50年生的恢复系列及对照样地(160~200 a)各设标准地5个,每个标准地面积为20 m ×20 m,每个标准地内设置4个10 m ×10 m的乔木和灌木样方,10个1 m ×1 m的草本样方。对乔木进行每木检尺,坐标定位,记录:高度、枝下高、冠层厚度、胸径、冠幅、林分郁闭度以及群落的发育程度;灌木和草本高度、盖度、种类、株数;海拔、坡向、坡度、坡位等生境因子。考虑到层间植物种类较少,将其以灌木计算。野外样方调查分别在2004年7—9月和2005年7—9月进行。
2.2 重要值计算方法(郝占庆等,2002b)乔木树种的重要值=(相对多度+相对显著度+相对频度)/3;
灌木(草本植物)的重要值=(相对高度+相对盖度+相对频度)/3。
2.3 α多样性测度方法选用Shannon指数H′来讨论群落的物种多样性,对群落的物种丰富度以物种数S来反映(郝占庆等,2002a;2002b):
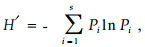
|
式中:Pi为种i的相对重要值,S为所计算的样地中的物种数目。群落的均匀度J的计算公式(郝占庆等,2002a)为
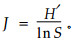
|
采用Simpson优势度指数D作为群落优势度测度的指标,其表达式为
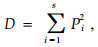
|
这里,多样性、均匀度与优势度的计算以重要值作为数量指标(彭少麟, 1996; 郝占庆等,2002a)。
2.4 β多样性测度方法Bray-Curtis指数CN的计算方法为
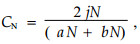
|
式中: aN为样地A的物种数目,bN为样地B的物种数目,jN为样地A(jNa)和样地B(jNb)共有种中个体数目较小者之和,以重要值作为测度指标。该指数是在Sorenson指数的基础上形成的(马克平等,1995; 高贤明等,1998)。
2.5 统计分析采用统计软件SPSS for windows 12.0中的双因素方差分析检验不同恢复系列和海拔梯度间的差异。用t检验分析不同生长型间的差异显著性。
3 结果与分析 3.1 不同恢复系列群落的物种组成在米亚罗地区海拔3 100~3 500 m的阴坡、半阴坡的箭竹红桦林和海拔3 300~3 600 m的阴坡、半阴坡的藓类红桦林中,所调查到的50个20 m×20 m样方共记录维管植物167种,隶属于44科117属。不同恢复系列,群落物种种类差别不大,但随着森林的恢复,优势物种的重要值不断发生变化。次生林乔木层树种主要由红桦、岷江冷杉以及紫果云杉(Picea purpurea)组成; 陕甘花楸(Sorbus koehneana)、康定野樱桃(Prunus tatsienensis)以及槭树(Acer spp.)主要出现在箭竹红桦林中; 而湖北花楸(Sorbus hupehensis)、微毛野樱桃(Prunus pilosiuscula)主要出现在藓类红桦林中。箭竹红桦林和藓类红桦林灌木层除了峨眉蔷薇(Rosa omeiensis)、红脉忍冬(Lonicera nervosa)、紫花卫矛(Euonymus porphyreus)等主要的共有种以外,2种次生林还有其各自的优势物种,在箭竹红桦林中有箭竹、木帚栒子(Cotoneaster dielsianus)、鞘柄菝葜(Smilax stans)、鲜黄小檗(Berberis diaphana)、五加等,另外还出现了五角槭(Acer mono)、疏花槭(Acer laxiflorum)、青荚叶(Helwingia japonica)等一些暖温性植物;在藓类红桦林中,主要有冰川茶藨子(Ribes glaciale)、散生栒子(Cotoneaster davaricatus)等。草本层除了蝎子草、鹿药、细辛、鹿尔韭、耳翼蟹甲草(Parasenecio otopteryx)、掌裂蟹甲草(Parasenecio palmatisectus)以外,在箭竹红桦林中还有扭柄花、管花鹿药(Smilacina henryi)、麦冬(Ophiopogon japonicus)、鬼灯擎(Rodgersia aesculifolia)、血见愁老鹳草(Geranium henryi)、四叶葎(Galium bungei);在藓类红桦林中,还有山酢浆草、猪秧秧(Galium spp.)、鸢尾(Iris spp.)、毛连菜(Picris hieracioides)等。原始暗针叶老龄林乔木层优势树种为岷江冷杉,灌木层除了冰川茶藨子、红脉忍冬、麻核栒子(Cotoneaster foveolatus)等共有种以外,在箭竹原始暗针叶老龄林中主要以箭竹占优势;草本层主要有蝎子草、高山露珠草等。阳生树种红桦寿命长达160年,在短时间内不会退出林冠层,在群落中仍发挥着建群种的作用,耐荫或中生性物种相继在群落中出现。
3.2 不同恢复系列群落的α多样性图 1可以看出,不同恢复系列次生林随着林龄增大,群落物种丰富度显著增加(表 1),最大值都出现在50年生的次生林中,50年生的箭竹红桦林物种丰富度为77,50年生的藓类红桦林为67,30、40年生物种丰富度上升最快,而后缓慢上升到一个峰值,到原始暗针叶老龄林时维持在一定水平。不同恢复系列次生林群落生长型之间物种丰富度差异极显著(表 2)。不同生长型物种丰富度为草本层>灌木层>乔木层,反映了群落内物种多样性空间分布格局。不同生长型物种丰富度最大值出现的时间不尽相同,乔木层物种丰富度最大值分别出现在40年生的箭竹红桦林和30年生的藓类红桦林,灌木层分别出现在50年生的箭竹红桦林和30年生的藓类红桦林,草本层都出现在50年生的次生林中。

|
图 1 不同恢复系列群落物种丰富度 Fig. 1 Species richness of various restoration series A:箭竹红桦林和原始暗针叶老龄林Bamboo-birch forests and old-growth bamboo-dark coniferous forests; B:藓类红桦林和原始暗针叶老龄林Moss-birch forests and old-growth moss-dark coniferous forests.下同The same below. |
|
|
|
|

群落物种均匀度是指群落中各个物种的多度或重要值的均匀程度。图 2可以看出,群落均匀度在40年生时达最大值。对箭竹红桦林,此后群落均匀度基本稳定在0.870 0左右。藓类红桦林在40年生时达最大值0.901 8,到50年生时又有所下降,到藓类原始暗针叶老龄林略有增大。不同恢复系列均匀度指数差异显著,2种森林类型间差异不显著(表 1)。t检验表明,乔木与灌木层以及乔木与草本层之间均匀度指数差异显著,而灌木与草本层间差异不显著(表 2)。

|
图 2 不同恢复系列群落均匀度 Fig. 2 Evenness of various restoration series |
Shannon指数随次生林恢复时间的增加呈增大的趋势(图 3),箭竹红桦林不同恢复系列最大值出现在50年生的林分中,为3.796 7;藓类红桦林最大值出现在30年生的林分中,为3. 654 3;箭竹和藓类原始暗针叶老龄林Shannon指数分别为3.563 0和3.401 0。不同恢复系列以及生长型之间Shannon指数差异显著,2种森林类型间差异不显著(表 1、2)。次生林Shannon指数为2.694 1~3.796 7,平均为3.473 2,老龄林为3.482 0,略高于次生林的平均值。Shannon指数的变化趋势与物种丰富度和均匀度大致相似。这可能是由于Shannon指数受物种丰富度和均匀度的双重影响有关。t检验表明,不同生长型之间Shannon指数差异显著(表 2),总体上来看,草本层>灌木层>乔木层。

|
图 3 不同恢复系列群落Shannon指数 Fig. 3 Shannon index of various restoration series |
群落优势度指数总体上呈先减小后增大的趋势,变化幅度相对较小(图 4)。不同恢复系列以及次生林类型间差异不显著(表 1)。乔木层与灌木层以及乔木层和草本层之间优势度差异显著,灌木层与草本层之间优势度差异不显著(表 2)。

|
图 4 不同恢复系列群落优势度 Fig. 4 Dominance index of various restoration series |
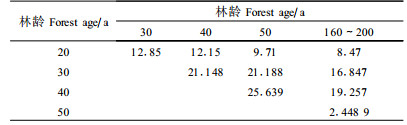
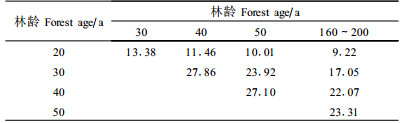
β多样性是群落多样性的重要内容。它可以定义为沿着某一环境梯度物种替代的程度或速率、物种周转率、生物变化速度等,β多样性还反映了不同群落间物种组成的差异。Bray-Curtis指数是一个应用广泛的指数,可用来测度群落或生境间的β多样性。Bray-Curtis指数值越大,说明群落间的共有种越多,物种组成越相似。从表 3和表 4可以看出,相邻恢复系列群落间的Bray-Curtis指数最大,说明群落间的相似性最大;恢复系列相距越远,Bray-Curtis指数越小,说明群落生境或物种组成差异随着林龄差异的增加而增大。随着林龄的增大,与原始暗针叶老龄林间的Bray-Curtis指数也有增大的趋势。
|
|
|
|
由于森林采伐破坏了原有生境, 群落物种组成和群落结构发生了一系列的变化。随着森林恢复, 红桦在不同恢复演替系列中成为优势种群。岷江冷杉也慢慢进入主林层, 在不同的恢复系列群落中出现了一些早期伴生种。箭竹红桦林由于分布海拔比藓类红桦林低, 不同恢复系列中, 乔木层伴生树种主要为一些半耐阴或喜暖温的树种, 如康定野樱桃和陕甘花楸, 而海拔较高些的藓类红桦林的伴生种主要为一些耐寒或较耐阴的树种, 如微毛野樱桃和湖北花楸。同一森林类型的不同恢复系列,群落物种种类差别不大,但随着森林的恢复,优势物种的重要值不断发生变化。
物种丰富度随着森林的恢复而逐渐增大, 在30、40年物种丰富度上升最快,而后缓慢上升到1个峰值,到原始暗针叶林老龄林时维持在一定水平。群落不同生长型物种丰富度具有相似的变化规律,这与其他学者的研究结果相似(贺金生等, 1997;温远光,1998)。物种丰富度随次生林的恢复不断增加,这可能是由于森林采伐后,土壤种子库中仍有大量的原有植物的种子,为群落的恢复提供了大量的种源。随着恢复时间的增加,林下幼苗和幼树生长发育进入林冠层,因此物种丰富度逐渐增大。恢复到30年后,许多先锋种仍然存在,而此时群落的生境为许多耐阴性树种提供了适生的条件,从而导致了这一时期物种丰富度急剧上升,到原始暗针叶老龄林阶段,许多阳生性物种已从群落中消失,岷江冷杉成为顶极优势种,群落中的物种丰富度维持在一定水平。
群落优势度指数与均匀度指数、Shannon指数以及物种丰富度呈相反的变化, 总体上呈先减小后增大的趋势。优势度指数减小的原因可能是由于森林的恢复,岷江冷杉、花楸、樱桃、槭树等逐渐进入乔木层,物种多样性增加,一定程度上削弱了早期占绝对优势的红桦的地位。到原始暗针叶老龄林时,其他树种逐渐消失,由岷江冷杉占绝对的优势,从而使得优势度增大;另外, 藓类红桦林乔木层优势度下降到最小值的时间箭竹红桦林要早,这可能是由于藓类红桦林的分布海拔比箭竹红桦林相对高些,更接近更高海拔处暗针叶林的保留林带,相对充足的种源使得藓类红桦林的岷江冷杉更新比箭竹红桦林快,从而相对较早地削弱了由单一优势树种红桦组成的群落,导致乔木层优势度降低,随着天然更新的进行,岷江冷杉在群落中的地位逐渐增大,形成与红桦抗衡的格局,最后基本取代了红桦,岷江冷杉成为顶级优势种,乔木层优势度降低经历由小到大的发展过程。
本文主要在川西亚高山原生暗针叶林海拔3 100~3 600 m的分布区内,对箭竹冷杉林和藓类冷杉林皆伐后形成的不同恢复演替阶段次生林群落物种组成和多样性的动态进行研究。森林皆伐20~30年后,一些强阳性物种比如悬钩子(Rubus spp.)逐渐消失,先锋树种、半耐阴或中生性树种如红桦、忍冬、花楸(Sorbus spp.)、茶藨子等侵入,同时冷、云杉幼苗、幼树也出现,这与该地区暗针叶林采伐迹地早期植被自然演替以及人工更新恢复过程中物种种类的变化一致(史立新等,1988;包维楷等,2002)。次生林物种多样性总体上随着森林的恢复呈增加的趋势,这与该地区人工针叶林恢复过程中物种多样性的变化有类似的结果(吴彦等,2001;赵常明等,2002)。恢复到暗针叶原始老龄林(160~200年)时物种多样性维持一定水平的发展。另外,β多样性是一个相对值,因此,运用Bray-Curtis指数来测度群落间β多样性的变化更重要的是找出不同恢复系列群落相似性的变化趋势。以重要值作为指标计算该指数较客观地反映了次生林恢复过程中群落间β多样性的动态。在本研究中, 随着林龄的增大,与原始暗针叶老龄林间的Bray-Curtis指数也有增大的趋势。这一结论与温远光在研究广西大明山中山植被恢复过程中群落间的变化相似(温远光,1998)。
本文中不同恢复系列物种组成和多样性的这种动态变化初步反映了生态系统恢复过程中群落环境的变化和生物多样性对这种变化的响应过程。50年相对于漫长的森林恢复演替过程还只能说是森林恢复初期或初中期阶段,全面揭示从森林皆伐后开始的次生林恢复演替到地带性顶级群落的整个恢复过程中物种多样性的变化规律还有待进一步深入研究。
包维楷, 张镱锂, 王乾, 等. 2002. 青藏高原东部采伐迹地早期人工重建序列梯度上植物多样性的变化. 植物生态学报, 26(3): 330-338. DOI:10.3321/j.issn:1005-264X.2002.03.012 |
高贤明, 马克平, 黄建辉, 等. 1998. 北京东灵山地区植物群落多样性的研究:Ⅺ.山地草甸β多样性. 生态学报, 18(1): 24-32. DOI:10.3321/j.issn:1000-0933.1998.01.004 |
郝占庆, 于德永, 杨晓明, 等. 2002a. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 13(7): 785-789. |
郝占庆, 郭水良, 曹同. 2002b. 长白山植物多样性及其格局. 沈阳: 辽宁科学技术出版社.
|
贺金生, 陈伟烈. 1997. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 17(1): 91-99. DOI:10.3321/j.issn:1000-0933.1997.01.014 |
蒋有绪. 1963a. 川西米亚罗高山暗针叶林的群落特点及其分类原则. 植物生态学与地植物学丛刊, 1(1): 42-50. |
蒋有绪. 1963b. 川西米亚罗、马尔康高山林区生境类型的初步研究. 林业科学, 8(4): 321-335. |
林开敏, 俞新妥, 黄宝龙, 等. 2001. 杉木人工林林下植物物种多样性的动态特征. 应用与环境生物学报, 7(1): 13-19. DOI:10.3321/j.issn:1006-687X.2001.01.003 |
刘庆, 吴彦, 何海. 2001. 中国西南亚高山针叶林的生态学问题. 世界科技研究与发展, 23(2): 63-69. DOI:10.3969/j.issn.1006-6055.2001.02.016 |
马克平, 刘灿然, 刘玉明. 1995. 生物群落多样性的测度方法:Ⅱ. β多样性的测度方法. 生物多样性, 3(1): 38-43. DOI:10.3321/j.issn:1005-0094.1995.01.007 |
彭少麟. 1996. 南亚热带森林群落动态学. 北京: 科学出版社.
|
石胜友, 杨季冬, 王周平, 等. 2002. 缙云山风灾迹地人工混交林生态恢复过程中物种多样性研究. 生物多样性, 10(3): 274-279. DOI:10.3321/j.issn:1005-0094.2002.03.004 |
史立新, 王金锡, 宿以明, 等. 1988. 川西米亚罗地区暗针叶林采伐迹地早期植被演替过程的研究. 植物生态学与地植物学学报, 12(4): 306-313. |
史作民, 程瑞梅, 刘世荣, 等. 2002. 宝天曼植物群落物种多样性研究. 林业科学, 38(6): 17-23. DOI:10.3321/j.issn:1001-7488.2002.06.004 |
温远光. 1998. 常绿阔叶林退化生态系统恢复过程物种多样性的发展趋势与速率. 广西农业大学学报, 17(2): 93-106. |
吴彦, 刘庆, 乔永康, 等. 2001. 亚高山针叶林不同恢复阶段群落物种多样性变化及其对土壤理化性质的影响. 植物生态学报, 25(6): 648-655. DOI:10.3321/j.issn:1005-264X.2001.06.002 |
阎海平, 谭笑, 孙向阳, 等. 2001. 北京西山人工林群落物种多样性的研究. 北京林业大学学报, 23(2): 16-19. DOI:10.3321/j.issn:1000-1522.2001.02.004 |
杨万勤, 王开运, Kellomäiki S. 2003. 生物多样性与针叶林生态系统过程的关系. 世界科技研究与发展, 25(5): 47-55. DOI:10.3969/j.issn.1006-6055.2003.05.008 |
张继义, 赵哈林, 张铜会, 等. 2004. 科尔沁沙地植被恢复系列上群落演替与物种多样性的恢复动态. 植物生态学报, 28(1): 86-92. DOI:10.3321/j.issn:1005-264X.2004.01.013 |
张远东, 赵常明, 刘世荣. 2005a. 川西米亚罗林区森林恢复的影响因子分析. 林业科学, 41(4): 189-193. |
张远东, 刘世荣, 马姜明, 等. 2005b. 川西亚高山桦木林的林地水文效应. 生态学报, 25(11): 2939-2946. |
赵常明, 陈庆恒, 乔永康, 等. 2002. 青藏东缘岷江上游亚高山针叶林人工恢复过程中物种多样性动态. 植物生态学报, 26(增刊): 20-29. |
Grime J P. 1997. Biodiversity and ecosystem function: the debate deepens. Science, 277: l260-1261. DOI:10.1126/science.277.5323.260 |
Guariguata M R, Chazdon R L, Denslow J S, et al. 1997. Structure and floristics of secondary and old-growth forest stands in lowland Costa Rica. Plant Ecology, 132: 107-120. DOI:10.1023/A:1009726421352 |
Ishida H, Hattori T, Takeda Y. 2005. Comparison of species composition and richness between primary and secondary lucidophyllous forests in two altitudinal zones of Tsushima Island, Japan. Forest Ecology and Management, 213: 273-287. DOI:10.1016/j.foreco.2005.03.046 |
Ruiz-Jaen M C, Aide T M. 2005. Restoration success: how is it being measured?. Restoration Ecology, 13(3): 569-577. |
Tilman D, Downing J A. 1994. Biodiversity and stability in grassland. Nature, 3: 363-365. |
Vázquez G J A, Givnish T J. 1998. Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra de Manantlán. Journal of Ecology, 86: 999-1020. DOI:10.1046/j.1365-2745.1998.00325.x |
Young T P. 2000. Restoration ecology and conservation biology. Biological Conservation, 92: 73-83. DOI:10.1016/S0006-3207(99)00057-9 |
Zerbe S, Kreyer D. 2006. Introduction to special section on "ecosystem restoration and biodiversity: how to assess and measure biological diversity?". Restoration Ecology, 14(1): 103-104. |