 2007, Vol. 43
2007, Vol. 43文章信息
- 何斌, 贾黎明, 金大刚, 秦武明.
- He Bin, Jia Liming, Jin Dagang, Qin Wuming.
- 广西南宁马占相思人工林土壤肥力变化的研究
- Studies on Soil Fertility Change in Acacia mangium Plantation in Nanning, Guangxi
- 林业科学, 2007, 43(5): 10-16.
- Scientia Silvae Sinicae, 2007, 43(5): 10-16.
-
文章历史
- 收稿日期:2006-03-01
-
作者相关文章


2. 北京林业大学 北京 100083;
3. 广西林业局 南宁 530022
2. Beijing Forest University Beijing 100083;
3. Guangxi Forestry Department Nanning 530022
马占相思(Acacia mangium)原产于澳大利亚昆士兰北部沿海、巴布亚新几内亚等地, 具有干形通直、出材率高、适应性广、耐干旱瘠薄、病虫害少、固氮改良土壤、速生丰产、用途广泛等优点(潘志刚等, 1996)。马来西亚于1966年在沙巴开始引种, 其后东南亚、美国、非洲等地先后大量引种, 人工造林面积迅速扩展。中国于1979年开始引进, 并从20世纪80年代中期开始在热带、南亚热带地区的广东、广西和海南等省区大面积种植和发展, 成为当地荒山绿化的主要树种之一, 同时也是仅次于桉树(Eucalyptus)的短周期速丰林树种, 并取得了显著的生态和经济效益。随着马占相思人工林经营规模的不断扩大, 国内外对马占相思的相关研究也日益增多, 其中对马占相思人工林土壤肥力性状方面的研究已有报道(Fisher, 1995; Kadir et al., 2003; Mackensen et al., 2003;蒋云东等, 2000; 许松葵等, 2003; 薛立等, 2003), 但仅限为对马占相思某一生长发育阶段(或某一年龄阶段)土壤理化性状的研究。本研究通过对不同年龄阶段(4、7和11年生)马占相思人工林土壤水分-物理性质、化学性质和生物化学性质(土壤酶活性)进行较系统的研究, 以揭示马占相思人工林土壤肥力演变规律与趋势, 为维护、保持以至提高人工林土壤肥力, 促进短周期工业用材林的可持续发展提供科学依据。
1 试验地及林分概况试验地位于广西南宁市国有高峰林场界牌分场(108°21′ E, 22°58′ N), 属南亚热带季风气候, 年均气温21.8 ℃, 极端最高气温40 ℃, ≥10 ℃年积温7 200 ℃, 年均降水量1 350 mm, 降水多集中在5—9月, 相对湿度79%。标准地位于山坡中部, 海拔250 m, 坡度25~28 °, 坡向南偏东, 土壤类型为砂页岩发育形成的赤红壤, 土层厚度在80 cm以上, 腐殖质层厚度15~20 cm。
标准地前茬林分均为杉木(Cunninghamia lanceolata)纯林, 林下植被主要有黄茅草(Heteropogon ontomtus)、五节芒(Miscanthus floridulus)、路边青(Clerodendrum bungei)等, 分别于各自造林前1年的年底采伐, 经炼山整地后, 于次年4月份用马占相思实生苗定植, 调查时4年生马占相思人工林的林相整齐, 郁闭度为0.75, 保留密度1 140株·hm-2, 林分平均树高12.0 m, 平均胸径(带皮, 以下同)14.0 cm, 林下植被以黄茅草、五节芒、路边青等为优势种, 此外还少量分布有桃金娘(Rhodomyrtus tomentosa)、野牡丹(Medinilla condidum)、粗叶悬钩子(Rubus alceaefolius)等, 覆盖度约70%, 凋落物层厚度为2~3 cm;7年生马占相思人工林经间伐和林分自疏后林相较整齐, 郁闭度0.85, 保留密度930株·hm-2, 林分平均树高17.7 m, 平均胸径18.3 cm, 林下植被以毛桐(Mallotus barbatus) 、桃金娘、野牡丹、乌毛蕨(Blechnum orientale)等为优势种, 此外还分布较多的五节芒、黄茅草, 粗叶悬钩子等, 覆盖度约70%, 凋落物层厚度3 cm;11年生马占相思人工林经间伐和林分自疏后林相较整齐, 郁闭度为0.85, 保留密度775株·hm-2, 林分平均树高为19.0 m, 平均胸径为23.5 cm, 林下植被主要以潺稿树(Litsea glutinosa)、盐肤木(Rhus chinensis)、九节(Psychotria rubra)、狗脊(Woodwardia japonica)等为优势种, 此外还少量分布有铁芒箕(Dicranopteris linearis)、五节芒、粗叶悬钩子等, 覆盖度约70%, 凋落物层厚度为4~5 cm。
2 研究方法 2.1 标准地设置与土壤样品采集在经过全面实地调查并分析林分状况的基础上, 分别在4年生(幼龄林)、7年生(中龄林)和11年生(近熟林)马占相思人工林中选择邻近分布, 母岩一致, 海拔、坡向、坡位等立地条件基本相同, 整地、抚育、施肥等营林和管理措施也相同, 长势良好且较一致的林分, 按不同年龄阶段设置面积20 m×20 m的标准样地各3块(其中4年生作为固定样地, 在栽植马占相思人工林以前即原杉木林已设置采样点采集土壤样品)。在每个标准地内设置代表性采样点3个, 按0~20和20~40 cm分层采集土壤样品, 把相同标准地同一层次土壤按质量等比例混合, 供测定土壤理化性质和酶活性使用, 同时用环刀采集土壤样品, 供测定土壤水分-物理性质使用。土壤样品采集时间为2005年3月。
2.2 分析方法 2.2.1 土壤水分物理性质与团聚体组成土壤-水分物理性质用环刀法测定, 土壤团聚体组成采用机械筛分法测定(中国科学院南京土壤研究所, 1978)。文中的试验数据为3块标准地测定结果平均值, 以下同。
2.2.2 土壤化学性质pH值采用电位法测定; 有机质用重铬酸钾氧化-外加热法测定(姜培坤, 2005);全氮用浓H2SO4-HClO4消化法消化-氨气敏电极法测定(何斌等, 2002b); 水解氮用碱解扩散法测定; 全P用氢氧化钠碱熔-钼锑抗比色法测定; 有效磷用HCl-H2SO4浸提-钼锑抗比色法测定; 全钾用氢氧化钠碱熔-火焰光度计法测定; 速效钾用乙酸铵浸提-火焰光度计法测定; 土壤腐殖质组成用焦磷酸钠提取-重铬酸钾氧化法(中国科学院南京土壤研究所, 1978)。
2.2.3 土壤酶活性过氧化氢酶用J.C.Johnson和K.L.Temple法测定; 蛋白酶用G.Hoffmann和K.Teicher法测定; 脲酶用G.Hoffmann与K.Teicher法测定; 酸性磷酸酶用G.Hoffmann法测定(关松荫,1986)。
3 结果与分析 3.1 土壤结构状况土壤团聚体的大小、数量和稳定性, 决定着土壤孔隙的大小和结构的稳定性, 影响着土壤的通气透水性和抗蚀性, 是土壤肥力的重要指标之一。表 1表明:4、7和11年生马占相思林0~20 cm土层的>5 mm的水稳性大团聚体含量比原杉木林分别增加2.79%、24.39%和51.14%, >0.25 mm的水稳性团聚体也分别增加了1.58%、5.34%和7.24%, 20~40 cm土层也有此趋势。而用来表征土壤结构稳定性的土壤结构破坏率则以11年生最小, 然后是7年生和4年生, 原杉木林最大, 20~ 40 cm土层也有相同趋势。土壤结构破坏率的降低意味着土壤水稳性团聚体数量提高, 土壤结构稳定性好, 遇雨时土体不易崩解, 抗蚀性能较强, 有利于水土保持。由此可见, 营造马占相思林后, 由于林分生长快, 枯枝落叶较多(据笔者测定, 4、7和11年生马占相思林当年凋落物量分别达到6.88、7.62和7.04 t·hm-2, 其中比较容易分解的落叶均占80%以上), 有利于土壤有机质含量的提高, 对土壤良好结构的形成与稳定有明显的促进作用, 土壤水稳性团聚体的数量和质量均有所增加和提高。
|
|
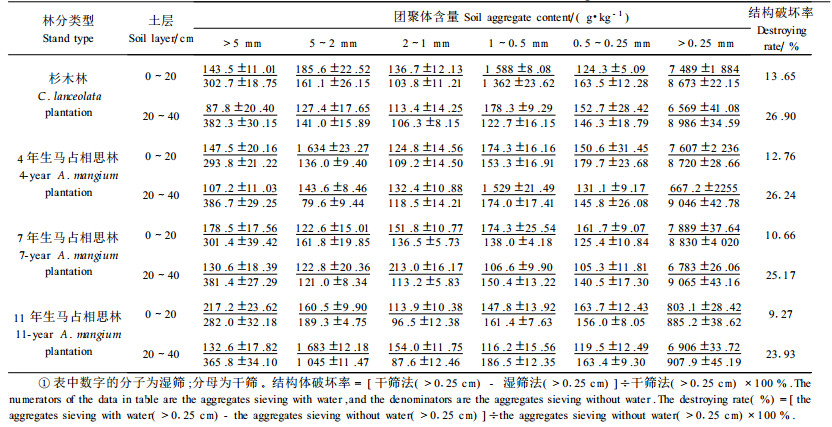
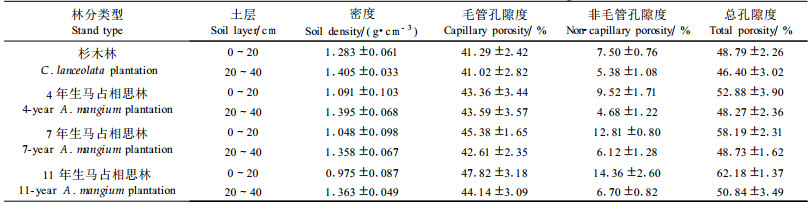
土壤密度和孔隙状况是土体构造虚实松紧的反映, 影响着土壤通气性、透水性和根系的伸展, 是表征土壤肥力的重要指标。表 2表明, 与原杉木林相比, 4、7和11年生马占相思林0~20 c m土层的密度分别下降了14.96%、18.32%和26.34%, 总孔隙度则分别增加8.38%和19.27%、27.44%, 其中毛管孔隙度分别增加5.01%、9.90%和15.81%, 非毛管孔隙分别增加2 6.93%、70.8%和91.47%, 20~40 cm土层也有相同趋势; 土壤团聚体、孔隙状况和颗粒组成综合反映的土壤持水供水特性中, 马占相思林地0~20 cm土层最大持水量、毛管持水量、田间持水量和自然含水量也均随林龄增长呈现增加趋势(表 3)。这一结果表明:随着马占相思的生长, 产生大量凋落物, 其分解后提高了土壤有机质含量, 加上马占相思发达根系(生物量占全树18.6%以上, 并主要分布于0~40 cm)(秦武明等, 2007), 改良了土壤结构, 减弱了地表径流和土壤水分的蒸发损失, 并使土体构造相对疏松, 通气性能更好, 容蓄能力或涵养水源能力也增强, 有利于林地水土保持和林木的生长。
|
|
|
|
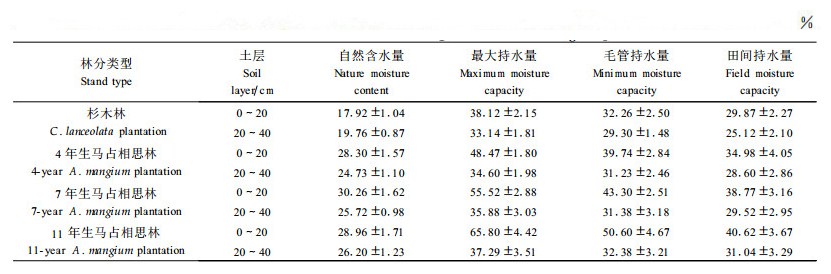
土壤腐殖质是土壤有机质的主体, 它的组成和性质既反映了土壤的形成条件(包括母质、植被和水热条件), 同时也反映了土壤肥力的高低变化。分析结果表明(表 4):与原杉木林相比, 4、7和11年生0~20 cm土层有机碳含量分别增加了31.08%、56.79%和73.38%;而从土壤胡敏酸碳(Humic acid C, HA)和富里酸碳(Fulvic acid C, FA)含量来看, 不论是0~20 cm土层, 还是20~40 cm土层, 它们的变化规律与有机碳含量相一致, 均为11年生>7年生>4年生>原杉木林。可见, 马占相思林的生长过程对土壤腐殖质(包括胡敏酸和富里酸)的积累有明显的促进作用。回归分析表明:马占相思林土壤胡敏酸碳和富里酸碳与有机碳的相关系数分别为0.968和0.971(n=18), 均达到极显著相关(P<0.01), 表明它们之间的关系极为密切。而不同年龄阶段马占相思人工林土壤腐殖质组成均以富里酸为主, HA/FA值均在0.60以下, 可见这些腐殖质的腐殖化程度均较弱, 均属于富里酸型, 土壤中合成的胡敏酸量少, 复杂程度较低, 但却有利于富里酸的形成和积累, 使后者成为主体, 这与相同地带性土壤的腐殖质组成特性相一致(李忠佩等, 1998)。
|
|
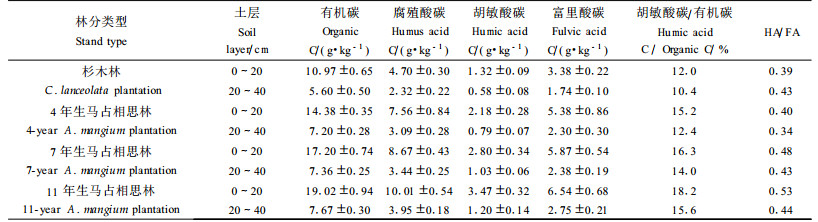
从表 4还可以看出, 随着马占相思的生长, 林地土壤HA/FA值呈现增大趋势, 其中以0~20 cm土层尤为明显, 而衡量土壤腐殖质品质优劣的主要标志之一的土壤腐殖化度, 即土壤胡敏酸占土壤全碳量的百分比(林明海等, 1982), 也有相同的变化趋势。可见, 随着马占相思的生长, 不但在数量上提高了土壤有机质含量, 而且提高了土壤的腐殖质聚合程度, 腐殖质的分子量增大, 复杂程度增加, 因而有利于土壤腐殖质品质的改善。
3.4 土壤养分状况在森林形成和生长的不同阶段, 受到林分结构、植被类型、盖度, 尤其是枯落物数量和特征等多方面影响, 森林土壤养分状况差异较大。从表 5可见, 与原杉木林相比, 4、7和11年生马占相思林0~20 cm土层全氮、全磷含量随着林分生长呈明显的增大趋势, 其中以全氮含量的增大趋势最明显, 分别增加了50.54%、107.20%和139.85%, 而反映土壤有机质矿质化和腐殖化程度的土壤有机质的C/N比值则随林龄的增加而减小, 一方面说明马占相思的生长过程对土壤有机质、全氮和全磷的生物积累均有明显的促进作用, 同时也显示出作为豆科植物的马占相思具有较强的固氮改土能力。表 5同时表明:除4年生马占相思林0~20 cm土层碱解氮含量与原杉木林基本一致外, 7年生和11年生马占相思林0~20 cm土层碱解氮、速效磷含量基本上呈现随林龄增长而增加的趋势, 可见, 马占相思的生长过程对土壤碱解氮和速效磷的生物积累均有明显的促进作用。而与原杉木林相比, 除4年生马占相思林地0~20 cm土层速效钾含量略为增加外, 其他2个林龄0~20 cm土层速效钾含量呈现降低的趋势, 与土壤其他养分含量的变化趋势恰好相反, 这可能是由于随着马占相思林分的生长, 林地植被对土壤钾素需求增大, 而林地土壤速效钾含量本来就比较低, 虽然可以通过凋落物的分解为林地积累一定量钾元素, 但仍然较难满足林木生长的需求, 加上钾元素是以钾离子方式存在于土壤中, 容易随雨水流失, 从而出现土壤速效钾含量随林木的生长而减少的现象。
|
|
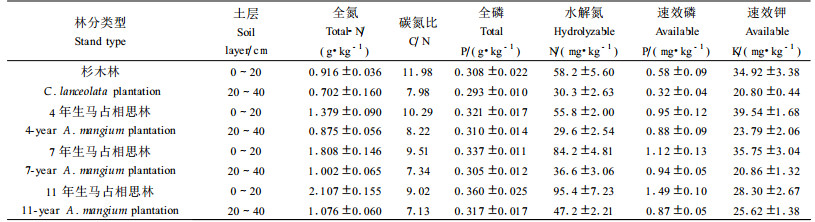
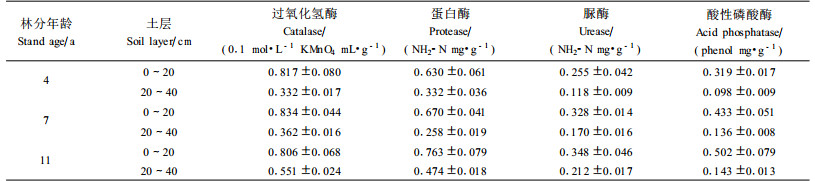
土壤酶是土壤的重要组成成分之一, 土壤中一切复杂的生物化学过程都是在酶的参与下进行的, 因此酶活性的高低, 不仅可以反映土壤生物化学过程的强度和方向, 而且还能客观反映土壤碳、氮、磷等的动态变化, 并与土壤主要肥力因素存在密切的关系(Peruci et al., 1984; 何斌等, 2002a; 孙启武等, 2003; 安韶山等, 2006)。不同年龄阶段马占相思人工林林地土壤酶活性及林地土壤酶活性与土壤养分间的相关性见表 6、7。
|
|
|
|
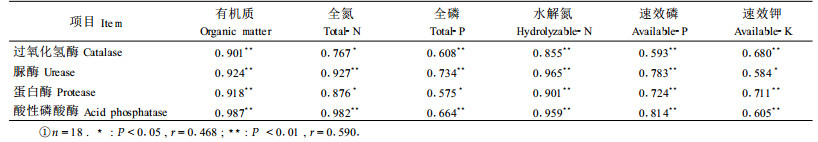
过氧化氢酶参与土壤中物质和能量转化, 具有酶促过氧化氢分解为水和氧分子,分解土壤中对植物有害的过氧化氢的作用, 在一定程度上反映土壤中腐殖质的再合成强度。研究表明:马占相思林地土壤过氧化氢酶在剖面的垂直分布规律与土壤有机质相一致, 并随林龄的增加呈增强趋势,过氧化氢酶不仅与有机质呈极显著正相关, 也与全氮、全磷和速效养分呈极显著正相关。
脲酶能分解有机物, 促进其水解生成氨和CO2, 其中氨是林木氮素营养的直接来源, 其活性可以用来表示土壤氮素状况。无论是0~20 cm土层还是20~40 cm土层, 脲酶活性均随马占相思林龄的增加而明显增强, 并与土壤各养分呈极显著正相关, 其中与水解氮和全氮的相关性最显著, 可见马占相思的生长有利于脲酶酶促反应的发生及进行, 有利于土壤中速效氮的转化, 也是其具有固氮改土作用的重要因素。
蛋白酶能酶促土壤中植物残体和微生物体的蛋白质水解成肽或氨基酸, 与脲酶一起参与土壤中的氮素转化, 对土壤生命和植物生长起着重要作用。研究结果显示:马占相思人工林0~2 0 cm土层蛋白酶活性也随马占相思林龄的增加而增强, 并与各土壤养分呈极显著或显著相关, 这与脲酶基本一致。
南方红壤中的磷酸酶以酸性磷酸酶为主, 它能促进土壤有机磷化合物的水解, 使之转化为植物可吸收利用的无机磷, 其活性反映土壤供应有效磷的潜在能力(何斌等, 2004)。0~20及20~40 cm土层的酸性磷酸酶活性与脲酶活性都有着相同的变化趋势, 与各土壤养分呈极显著相关, 其中与土壤有机质相关性最显著, 其次是全氮、水解氮和速效磷。因此, 其活性的强弱对马占相思林地土壤中磷素的供应也十分重要。
由此可见, 马占相思人工林3个不同年龄阶段的土壤酶活性, 不论是氧化还原酶的过氧化氢酶, 还是水解酶的脲酶、蛋白酶和酸性磷酸酶, 其活性基本上呈现11年生>7年生>4年生, 即随林龄的增加土壤酶活性增强, 这既体现出土壤酶活性与林地植物现存量或凋落物的关系, 即单位面积植物产量或凋落物量越多, 土壤酶活性越高(Sinsabaugh et al., 1993; Johansson, 1995; 张银龙等, 1999; 何斌等, 2002b), 同时也说明马占相思的生长有利于酶促作用的发生, 增强土壤酶活性。而4种土壤酶活性与土壤有机质均存在极显著正相关, 更进一步说明土壤有机质不但在很大程度上影响土壤的理化特性和水分特性,同时也是土壤酶活性的主要来源和储藏场所。
4 结论与讨论与原杉木林相比, 马占相思人工林0~20 cm土层的水稳性团聚体含量、非毛管孔隙度、毛管孔隙度及总孔隙度均有所提高, 并随马占相思林龄的增长而呈增大趋势, 土壤最大持水量、田间持水量及毛管持水量也呈现相同的变化趋势, 而表征土壤团聚体稳定性的结构体破坏率和土壤密度则呈明显下降趋势。表明栽植马占相思人工林有利于土壤良好结构的形成与稳定, 改善土壤-水分物理性质。
随林龄的增加, 林地土壤有机质、全N、全P和速效养分含量呈现增大趋势, 其中以全N的增大趋势最明显。与此同时, 作为衡量土壤腐殖质品质优劣的主要标志的土壤胡敏酸与富里酸之比(HA/FA)和土壤腐殖化度也随着马占相思林龄的增大而提高, 说明马占相思林生长过程不但在数量上提高了土壤腐殖质含量, 而且也改善土壤腐殖质的品质, 土壤的团粒化作用增强, 更有利于林木的生长发育。
马占相思林地土壤酶活性, 随林龄的增加而发生变化, 不论是氧化还原酶中的过氧化氢酶, 还是水解酶中脲酶、蛋白酶和酸性磷酸酶, 除11年生0~20 cm土层过氧化氢酶比4年、7年生略低外, 均随林龄的增长而增强, 说明马占相思的生长对土壤酶活性的提高有明显的促进作用。而马占相思林地土壤酶活性与土壤主要肥力因素有密切的关系, 表明土壤酶活性也可作为评价马占相思林地土壤肥力的指标之一。
森林的形成与生长过程, 也是森林与土壤相互影响和相互作用的过程, 森林群落的变化也总是与土壤性质的演化相关联, 土壤性质的变化会导致植被的变化, 而植被变化的同时也影响着土壤发育(张庆费等, 1999; 何斌等, 2002b; 焦如珍等, 2005)。在人工林的形成和生长过程的早期, 由于炼山、整地和植树作业, 生态系统原有植被和地表凋落物大部分被清除, 迹地裸露, 破坏了表土的物理性质, 使林地遭受降雨直接打击和地表径流冲蚀, 土壤结构变差, 土壤养分有所流失, 导致林地地力衰退(张昌顺等, 2005)。因此, 在炼山后栽植马占相思及其生长的过程, 实际上可看作林地土壤肥力恢复、维持与提高的过程。而由枯枝落叶层和土壤有机质构成的森林土壤有机物和养分库是森林生态系统物质循环的基础, 同时也是森林土壤肥力得以恢复、维持和提高的物质条件。研究表明:原杉木林土壤较贫瘠, 土壤水分-物理性质和化学性质均较差, 其林下植被以耐干旱、瘠薄的黄茅草、五节芒等占绝对优势; 在栽植马占相思后的幼龄林阶段(林龄≤4年), 林分生物量较小, 但由于其落叶的高氮量和较大的凋叶量, 加上其具有比较发达且分布较浅的根系, 使林下土壤快速改良, 因此, 其土壤肥力与原杉木林相比呈上升趋势, 林下植被虽仍然以黄茅草、五节芒等草本植物占优势, 但已有较多的对土壤肥力要求较高的灌木, 如桃金娘、野牡丹等, 说明土壤肥力得到了一定的改善。随着林龄增加, 林分生物量增大, 凋落物明显增多, 促进土壤腐殖质的合成和分解, 加快林地土壤养分的循环和积累, 有利于土壤良好结构的形成与稳定, 并改善了土壤水分-物理性质, 林下植被优势种由4年生的黄茅草、五节芒, 逐步发展到潺稿树、盐肤木、九节、狗脊等对土壤肥力要求较高的植物, 指示林地土壤肥力水平较大程度提高。由此可见, 在较贫瘠的立地营造马占相思林, 有利于维护和提高土壤肥力, 防止地力衰退, 马占相思林不仅具有较高的经济效益, 其生态效益也十分明显。目前中国南方大规模及多代连栽杉木和桉树对林地生态环境产生了不良影响, 笔者认为:在适地适树的前提下, 通过合理轮植, 如栽培2~3代杉木或桉树后, 接着栽培1代马占相思林, 或与桉树进行混交, 从而改良杉木或桉树林地土壤, 恢复并提高土壤肥力, 有利于实现人工林经营的经济效益与生态效益并举的目标, 促进林业的可持续发展。
安韶山, 黄懿梅. 2006. 黄土丘陵区柠条林改良土壤作用的研究. 林业科学, 42(1): 71-75. |
关松荫. 1984. 土壤酶及其研究法. 北京: 农业出版社, 274-319.
|
何斌, 刘运华, 陆志科, 等. 2004. 肉桂人工林土壤速效养分与酶活性的季节变化. 经济林研究, 22(3): 1-4. DOI:10.3969/j.issn.1003-8981.2004.03.001 |
何斌, 温远光, 梁宏温, 等. 2002a. 英罗港红树植物群落不同演替阶段植物元素分布及其与土壤肥力的关系. 植物生态学报, 26(5): 518-524. |
何斌, 温远光, 袁霞, 等. 2002b. 广西英罗港不同红树植物群落土壤理化性质与酶活性的研究. 林业科学, 38(2): 21-26. |
姜培坤. 2005. 不同林分下土壤活性有机碳库研究. 林业科学, 41(1): 10-13. |
蒋云东, 匡玉兰, 李思广, 等. 2000. 云南热区几种人工林土壤变化研究. 土壤与环境, 9(2): 110-113. DOI:10.3969/j.issn.1674-5906.2000.02.007 |
焦如珍, 杨承栋, 孙启武, 等. 2005. 杉木人工林不同发育阶段土壤微生物数量及其生物量的变化. 林业科学, 41(6): 16-65. DOI:10.3321/j.issn:1001-7488.2005.06.003 |
李忠佩, 程励励, 林心雄. 1998. 红壤腐殖质组成变化特点及其与肥力演变的关系. 土壤, 34(1): 9-15. |
林明海, 赖庆旺. 1982. 不同熟化度红壤及红壤性水稻土的腐殖质组成及特性. 土壤学报, 19(3): 237-247. |
潘志刚, 冯水, 林鸿盛. 1996. 马占相思引种、生长及利用. 热带林业, 24(4): 144-152. |
秦武明, 何斌, 余浩光, 等. 2007. 马占相思人工林不同年龄阶段的生物生产力. 东北林业大学学报, 35(1): 22-24. DOI:10.3969/j.issn.1000-5382.2007.01.007 |
孙启武, 杨承栋, 焦如珍. 2003. 江西大岗山连栽杉木人工土壤性质的变化. 林业科学, 39(3): 1-5. DOI:10.3321/j.issn:1001-7488.2003.03.001 |
许松葵, 薛立, 杨鹏, 等. 2003. 马占相思等5种纯林林地土壤理化性质分析. 广东林业科技, 19(4): 27-29. DOI:10.3969/j.issn.1006-4427.2003.04.006 |
薛立, 邝立刚, 陈红跃, 等. 2003. 不同林分土壤养分、微生物与酶活性的研究. 土壤学报, 40(2): 280-285. DOI:10.3321/j.issn:0564-3929.2003.02.018 |
张昌顺, 李昆. 2005. 人工林地力的衰退与维护研究综述. 世界林业研究, 18(1): 17-21. DOI:10.3969/j.issn.1001-4241.2005.01.004 |
张庆费, 宋永昌, 由文辉. 1999. 浙江天童植物群落次生演替与土壤肥力的关系. 生态学报, 19(2): 174-178. DOI:10.3321/j.issn:1000-0933.1999.02.006 |
张银龙, 林鹏. 1999. 秋茄红树林土壤酶活性时空动态. 厦门大学学报:自然科学版, 38(1): 129-136. |
中国科学院南京土壤研究所. 1978. 土壤理化分析. 上海: 上海科学技术出版社.
|
Fisher R F. 1995. Amelioration of degraded rain forest soil by plantations of native trees. Soil Science Society of America Journal, 59(2): 544-549. DOI:10.2136/sssaj1995.03615995005900020039x |
Johansson M B. 1995. The chemical composition of needle and leaf litter from Scots pine, Norway spruce and white birch in Scandinavian forests. Forestry, 68: 49-62. DOI:10.1093/forestry/68.1.49 |
Kadir S, Sakurai K, Tanaka S, et al. 2003. Characteristics of Ultisols differing in wildfire history in South Sumatra, Indonesia Ⅱ.Dynamics of chemical properties. Soil Science and Plant Nutrition, 49(1): 1-7. DOI:10.1080/00380768.2003.10409972 |
Mackensen J, Folster H. 2000. Cost analysis for a sustainable nutrient management of fast growing tree plantations in East Kalimantan, Indonesia. Forest Ecology and Management, 131(1/3): 239-253. |
Peruci P, Scarponi L, Businelli M. 1984. Enzyme activities in clay loam soil amended with various crop residue. Plant and Soil, 81: 345-351. DOI:10.1007/BF02323049 |
Sinsabaugh R L, Antibus R K, Linkins A E, et al. 1993. Wood decomposition:nitrogen and phosphorus dynamics in relation to extracellular enzyme activity. Ecology, 74: 1586-1593. DOI:10.2307/1940086 |