 2007, Vol. 43
2007, Vol. 43文章信息
- 赵文飞, 王迎, 王华田, 亓立云, 董玉峰.
- Zhao Wenfei, Wang Ying, Wang Huatian, Qi Liyun, Dong Yufeng.
- 不同季节麻栎树干贮水量的动态变化
- Change of Quercus acutissima Trunk Pondage in Different Season
- 林业科学, 2007, 43(4): 115-120.
- Scientia Silvae Sinicae, 2007, 43(4): 115-120.
-
文章历史
- 收稿日期:2006-04-04
-
作者相关文章



2. 山东省泰山林业科学研究院 泰安 271000
2. Taishan Academy of Forestry Science Tai'an 271000
森林个体和群体耗水特性对区域水量平衡的影响是评价森林生态效应的重要依据。以往有关林木耗水的测定技术和计算方法已经取得较大进展,在单枝、叶片、单木、林分空间尺度上逐渐趋于成熟(刘奉觉等, 1997; 孙慧珍等, 2004),应用上述方法研究林分个体和群体的耗水性已经成为近年来林木水分生理生态和森林水文的研究热点。
近年来,欧美学者应用液流检测技术对欧洲赤松(Pinus sylvestris)、欧洲云杉(Picea abies)、北美黄杉(Pseudotsuga menziesii)等树种的蒸腾耗水特性进行了研究(Moren et al., 2000; Cermak et al., 1995; Alarcón et al., 2003; Granier, 1987),并对影响树木蒸腾作用的环境因子进行了相关分析。Marshall(1958)、Swanson(1994)和Wullschleger等(1998)对单木整株耗水量进行了精确的测定,推求了不同树种林木个体和群体的耗水量。我国李海涛等(1998)最早应用热扩散技术研究了五角枫(Acer mono)树干液流传输规律。孙鹏森(2000)、王华田等(2002a;2004)、马履一等(2002; 2003)对刺槐(Robinia pseudoacacia)、栓皮栎(Quercus variabilis)、油松(Pinus tabulaeformis)、侧柏(Platycladus orientalis)等树种树干边材液流传输的时空变化规律进行了研究,比较了不同树种的耗水性,对林木耗水的时间和空间局限性进行了评述,对单木到林分时空尺度耦合问题进行了有益的尝试和探讨。周国逸等(2002)观察到TDP探针对树干不同位点边材液流的响应不同。此外,茎流技术在我国黑河流域的二白杨(Populus gansuensis)农田防护林(常学向等, 2004)和宁南华北落叶松(Larix principi-rupprechtii) (熊伟等, 2003)人工林中也有应用。
本文利用德国Ecomatik公司生产的热扩散茎流探针(thermal dissipation probe)SF-G对泰山林业科学研究院林场麻栎(Quercus acutissima)人工林中标准地平均木于春末、深秋两季的连日树干液流进行连续观测。据此比较了春、秋季麻栎边材液流特征和树干不同高度边材液流变化规律,深入分析树干贮水季节变化及其对边材液流的影响,为准确评价人工林群体耗水性和森林水量平衡提供依据。
1 研究地概况泰山林业科学研究院林场位于泰山罗汉崖,36°15′ N,117°04′ E,海拔280~400 m,坡位为山坡中部,坡度15~25°。母质为片麻岩,土壤为粗骨质棕壤,土层厚30~40 cm。属半湿润大陆性季风气候,年平均气温12.1 ℃,极端最低气温-20.8 ℃,极端最高气温40.3 ℃,年日照时数2 450 h,多年平均降水量690.3 mm,≥10 ℃有效积温4 300 ℃,无霜期198 d。主要植被是由油松、赤松(Pinus densiflora)、侧柏、麻栎、栓皮栎、刺槐、五角枫、车梁木(Cornus walteri)等树种组成的人工林,下木和草本植物主要是黄栌(Cotinus coggygria)、黄连木(Pistacia chinensis)、黄荆(Vitex negundo)、胡枝子(Lespedeza bicolor)、扁担木(Grewia biloba)、白羊草(Bothriochloa ischaemum)、黄背草(Themeda triandra)、鬼针草(Bidens pilosa)等。
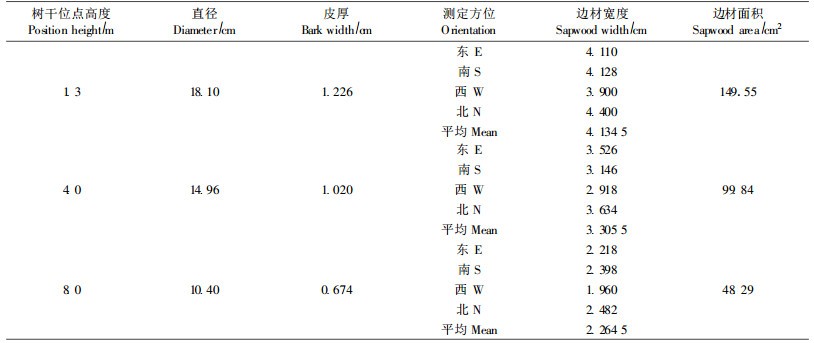
2 材料与方法对林分进行标准地调查,根据试验要求选择树干通直圆满、不偏心、不偏冠、测定部位无节疤或损伤的样木,实测样木生长参数为:树高11.3 m,胸径18.1 cm,冠幅7.1 m×4.2 m。利用生长锥测定边材宽度和心材宽度,计算边材面积(表 1)。在林分内部搭设钢架建筑平台(5.0 m×2.0 m×8.0 m),直达麻栎树木冠层,于平台4.0 m、8.0 m处铺放木板,用铁丝网在钢架周围架设围栏,分别在树干1.3 m、4.5 m、8.0 m处将麻栎外层栓皮刮掉,露出内层活树皮,安装茎流探针(TDP),方法参见王华田等(2002a)。分别于2005年5月30日— 6月3日、10月30日—11月4日2个阶段测定春、秋两季树干的茎流速率(春夏之交、深秋季节,是麻栎边材液流特征剧烈变化的时期),采集数据的时间间隔为30 min。树干水分流入量以1.3 m处液流通量计算。数据处理:利用Excel和SPSS10.0统计软件对观测数据进行统计分析、绘图和制表。
|
|
春、秋两季,麻栎边材液流存在明显差异。从图 1可以看出,春末麻栎进入迅速生长期,由于蒸腾作用逐渐增强,从而引起树体较高的边材液流速率;秋末麻栎停止生长,逐渐进入休眠期,蒸腾作用微弱,边材液流速率较小。
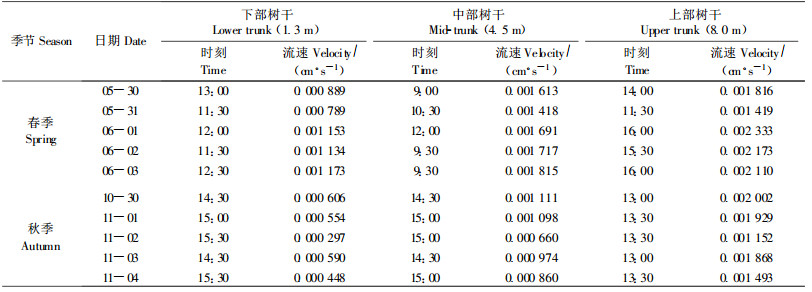
春季树干中部液流速率首先出现全天最大值,随后下部出现液流高峰,树干上部的液流高峰多出现在14:00—16:00。春季进入高峰期后持续时间较长,液流峰较宽,陡度小;而秋季则不同,树干上部首先在13:00—13:30出现边材液流高峰,树干中、下部位边材液流高峰要延迟60~120 min,秋季高峰期持续时间短,液流峰较尖,陡度大。春、秋两季边材液流高峰值差异表现为中、下部差异大,春季树干中、下部液流高峰约为秋季相同位点的1.5~2倍,树干上部液流峰值在春、秋季(未落叶前)表现不明显。春季树干不同位点高度液流启动时间彼此衔接,而秋季树干中、下位边材液流启动时间明显滞后于树干上部。

|
图 1 麻栎树干不同高度边材液流速率连日变化 Fig. 1 Diurnal sap flow velocity at different heights of Q.acutissima trunk |
|
|
边材液流通量能够直观地反映单株树木瞬态耗水特征。树干不同高度3个断面边材液流通量瞬态变化与液流速率变化不同。图 2表明,春季,树干中、下部位(4.5 m)边材液流通量峰值最高,数值相近;树干顶部(8.0 m)液流通量小,液流通量高峰期最短。秋季树干上、中、下3个位点边材液流速率相差较小。由于液流速率直接相关,所以秋季通过树干横断面的液流通量小于春季。整个树干(1.3~8.0 m)液流通量连日变化动态(规定树干内水分流入,液流通量值为正,水分流出为负),从图 2可以看出,春季,尽管麻栎蒸腾引起较高的边材液流速率,但树干多数时间是贮水阶段(液流量为正值),树干水分很少有亏损的时候;秋季则不同,全天树干水分流入、流出量持平(液流值正、负累积面积相抵),即树干液流通量0:00点之后为负值,并且逐渐减小,14:00—15:00之前树干内水分亏损,15:00之后是短时间的补充,且树体吸收的水分与损失的相差不大。

|
图 2 麻栎树干不同高度边材液流通量连日变化 Fig. 2 Diurnal sap flow density at different heights of Q.acutissima trunk |
对测定期间流经麻栎顶部和胸径断面的液流量进行每日累计(图 3)和连日累计(图 4)发现,5月30日—6月3日,流经样木胸径处和顶部的液流量分别为25 934.24 cm3,15 003.22 cm3,树干总盈余水分10 931.02 cm3,10月31日—11月4日,树干对应断面(1.3 m, 8.0 m)的液流量累计为9 800.47 cm3,8 933.836 cm3,树干水分总变化量为866.63 cm3。春季树干在正常的生长状况下,树干内部水分有增加的趋势(阴影部分高度逐渐增加),深秋麻栎树干水分保持相对稳定,尽管仍有短时蒸腾作用引起较少茎流,但树干总吸收水量与向冠层传输水量基本持平,总体来说,麻栎春天耗水累计曲线斜率大于秋季,耗水量远大于秋季。

|
图 3 麻栎树干边材液流每日累计量 Fig. 3 Accumulative total volume of sap flow at different trunk cross sections of Q.acutissima per day |

|
图 4 麻栎树干边材液流连日累计量 Fig. 4 Diurnal accumulative total volume of sap flow at different trunk cross sections of Q.acutissima |
从图 3可以更清楚地看到,春季树干每日累计吸水量(通过胸径处)远大于秋季,图中阴影部分表明了每日树干吸水和向冠层传输的水量差值。春季树体根系吸水大于冠层的蒸腾,树干水量有盈余,秋季上部蒸腾作用略大于根系吸水,阴影部分说明树干水量有增有减,但变化量不大。麻栎树干顶部与下部边材液流量变化关系如图 5所示,图中曲线表示树干相对于初始状态(春季初始状态日期为5月29日;秋季为10月30日)的贮水量变化,春季每日累计树干贮水量很难回到初始状态,相对每日吸收的水分,一般为盈余;秋季树干水分状态在白天和夜间可以2次恢复到初始状态,树干水分状况较稳定。

|
图 5 麻栎树干上部液流通量与下部液流通量关系图 Fig. 5 The relationship of the total sap flow flux of the upper and lower trunk of Q.acutissima |
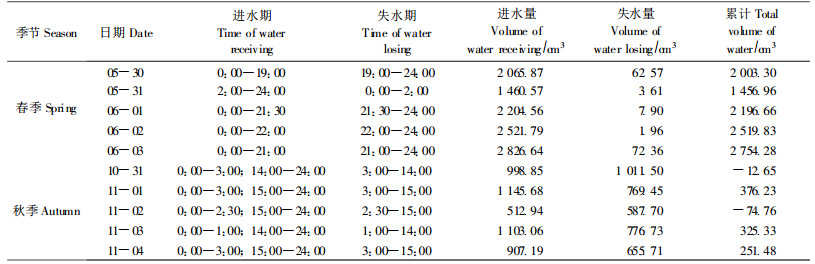
从表 3可以看出,春季麻栎树干进水相对时间较长,约为20 h,夜间树干向冠层传输水分的时间较短,约2~4 h。春季树干失水量很少,从而也在一定程度上说明冠层水分可以得到快速补充,从每日进、出树干水累计量看,春季树干存水较多。秋季,麻栎树干每天有2个进水时段,第1次一般在凌晨3:00以前,第2次在下午14:00以后,树干失水期也相对集中在凌晨至下午之间,此时树干水分向冠层流动较多;此外秋季麻栎树干进水与失水存在较小差值,树干水量变化平稳,树干存水较少。此种现象是否与麻栎休眠期为避免冬季冻害而采取的适应对策有关,有待进一步研究。
|
|
利用热扩散技术对春、秋两季麻栎边材液流的日变化、连日变化进行测定发现,其边材液流特征有明显的日节律和季节节律,春季麻栎有多个液流高峰出现,与核桃楸(Juglans mandshurica)(严昌荣等, 1999)相似,但液流速率与其他树种(常学向等, 2004; 熊伟等, 2003)存在差异,说明不同树种耗水特性有差别。秋季麻栎边材液流日变化多呈单峰曲线,液流速率小于春季,说明麻栎树干液流特征存在季节变异,作者认为可能受太阳辐射、空气和土壤温、湿度等环境因子年季节节律影响。
麻栎树干不同高度边材液流特征是:上部液流速率大,高峰期持续时间比中部和下部短,春、秋季节树干上、下部位最大液流速率均相差2~3倍。春季树干不同部位液流的启动时间早晚,情况比较复杂,主要取决于树体水分状况;秋季规律明显,树干中、下部明显滞后于上部,时间差在60~120 min不等。形成春季麻栎边材液流特征的主要原因可能是在土壤、树体水分储备足时,上部液流由于太阳辐射进行蒸腾作用,首先感受SPAC系统叶、气界面水势梯度变化,率先启动。如果夜间蒸腾停止时,树体正常水分尚未恢复,则是树干下部边材液流依然延续前期吸水过程,之后有可能短期吸水然后暂时停止(吸水充足),或感受由于冠层蒸腾引起树体自上而下的水势变化,继续流动。树干中部边材液流的特征可以视作树体上、下部位的衔接,亦反映树体水分流动、分配动态。树干不同高度边材液流速率的复杂变化也决定了通过树干不同高度横断面的液流通量。
对麻栎树干断面液流通量及日变化累计量进行的统计发现,树干木质部总是处在补水与耗水动态变化之中,但树体对水分的吸收和耗散不是等量的或者说是有延迟的。春季麻栎树干从基部上传的水分并未全部用于向冠层进行水分补充,而是有一定的盈余,贮存在树干内部,连日累计树干上、下部位断面的茎流通量差值,发现这种情况持续累计,说明林木树干类似容器有贮存水的特性,很好地证明了树体内部水容的存在,但这种效应必定最终因树种贮水特性所限,贮水过程不会持续下去,即树干贮水累计曲线(图 3)阴影部分高度不会持续增加,而是到达最大值后随即减小,并处于波动之中。王华田(2002b)认为水容的存在是树干不同部位之间液流速率产生差异的主要原因,对高大乔木而言,尽管根系的水容最高,由于树干是构成树体的主要器官,因此树干水容在整株水分传输过程中发挥的作用最大。秋季,树干贮水相对小,树干水分基本保持稳定。一方面说明,植物即将进入休眠期,树体各器官代谢节律降低,根系吸水减慢,叶部蒸腾作用变得愈加微弱;另一方面,说明气温变冷,土壤与根系的水分传输受到影响,水分运动变慢。这种现象在一定程度上体现了树木自身对环境因子的适应。水容是动态的、瞬时的,与树干含水量变化有关,受组织水势影响。目前,研究水容效应对植物耗水的调节,特别是高大乔木,困难很大,测定林木树干含水量的变化,通过比较树干不同高度断面茎流通量差值可以准确地进行估算,但对于树干水势的测定却遇到了难题,至今很难找到行之有效的方法,而这问题能否突破直接影响到对SPAC系统中植物体内水分传输途径及瞬态流特征等问题的认知程度,也因为如此,以前的众多研究把边材液流累计通量等同于树木的耗水量,从而忽略了高大树木的贮水特性——水容的作用。
常学向, 赵文智. 2004. 荒漠绿洲农田防护树种二白杨生长季节树干液流的变化. 生态学报, 24(7): 1437-1441. |
李海涛, 陈灵芝. 1998. 应用热脉冲技术对棘皮桦和五角枫树干液流的研究. 北京林业大学学报, 20(1): 1-6. DOI:10.3321/j.issn:1000-1522.1998.01.001 |
刘奉觉, 郑世锴, 巨关升. 1997. 树木蒸腾耗水测算技术的比较研究. 林业科学, 33(2): 117-126. DOI:10.3321/j.issn:1001-7488.1997.02.003 |
马履一, 王华田. 2002. 油松边材液流时空变化及其影响因子研究. 北京林业大学学报, 23(4): 23-37. DOI:10.3969/j.issn.1671-6116.2002.04.004 |
马履一, 王华田. 2003. 树木耗水性比较的研究. 北京林业大学学报, 24(2): 1-6. |
孙慧珍, 周晓峰, 康绍忠. 2004. 应用热技术研究树干液流进展. 应用生态学报, 15(6): 1074-1078. DOI:10.3321/j.issn:1001-9332.2004.06.032 |
孙鹏森. 2000.京北水源保护林树种不同尺度耗水特性及林分配置的研究.北京林业大学博士研究生学位论文
|
王华田, 马履一, 孙鹏森. 2002a. 油松侧柏深秋边材木质部液流变化规律的研究. 林业科学, 38(5): 31-37. |
王华田, 马履一. 2002b. 利用热扩散式边材液流探针(TDP)测定树木整株蒸腾耗水量的研究. 植物生态学报, 26(6): 661-667. |
王华田, 马履一, 徐军亮. 2004. 油松人工林SPAC水势梯度时空变化规律及其对边材液流传输的影响. 植物生态学报, 28(5): 637-643. DOI:10.3321/j.issn:1005-264X.2004.05.008 |
肖以华, 陈步峰, 陈嘉杰, 等. 2005. 马占相思树干液流的研究. 林业科学研究, 18(3): 331-335. DOI:10.3321/j.issn:1001-1498.2005.03.020 |
熊伟, 王彦辉, 徐德应. 2003. 宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应. 林业科学, 39(2): 1-7. |
严昌荣, Alec D, 韩兴国, 等. 1999. 北京山区落叶阔叶林中核桃楸在生长中期的树干液流研究. 生态学报, 19(6): 793-797. DOI:10.3321/j.issn:1000-0933.1999.06.005 |
周国逸, 黄志宏, Jim M, 等. 2002. 桉树人工林树液流动密度随边材径向深度的变化. 植物学报, 44(12): 1418-1424. DOI:10.3321/j.issn:1672-9072.2002.12.005 |
Alarcón J J, Domingo S R, Nicolás E, et al. 2003. Estimation of hydraulic conductance within field-grown apricot using sap flow measurements. Plant and Soil, 251: 125-135. DOI:10.1023/A:1022976110768 |
Cermak J, Ciencala E, Kucera J, et al. 1995. Individual variation of sap-flow rate in large pine and spruce trees and stand transpiration: a pilot study at the central NOPEX site. Journal of Hydrology, (168): 17-27. |
Granier A. 1987. Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements. Tree Physiology, 3: 309-319. DOI:10.1093/treephys/3.4.309 |
Marshall D C. 1958. Measurement of sap flow in conifers by heat transport. Plant Physiology, 33: 385-396. DOI:10.1104/pp.33.6.385 |
Moren A S, Lindroth A, FlowerEllis J, et al. 2000. Branch transpiration of pine and spruce to tree and canopy using needle biomass distributions. Tree, 14: 384-397. DOI:10.1007/PL00009774 |
Swanson R H. 1994. Water transpired by trees is indicated by heat pulse velocity. Agricultural Meteorology, 72: 113-132. DOI:10.1016/0168-1923(94)90094-9 |
Wullschleger S D, Meinzer F C, Vertessy R A. 1998. A review of whole-plant water use studies in trees. Tree Physiol, 18(8/9): 499-512. |