 2007, Vol. 43
2007, Vol. 43文章信息
- 范国强, 张胜, 翟晓巧, 刘飞, 董占强.
- Fan Guoqiang, Zhang Sheng, Zhai Xiaoqiao, Liu Fei, Dong Zhanqiang.
- 抗生素对泡桐丛枝病植原体和发病相关蛋白质的影响
- Effects of Antibiotics on the Paulownia Witches' Broom Phytoplasmas and Pathogenic Protein Related to Witches' Broom Symptom
- 林业科学, 2007, 43(3): 138-142.
- Scientia Silvae Sinicae, 2007, 43(3): 138-142.
-
文章历史
- 收稿日期:2006-09-29
-
作者相关文章


泡桐(Paulownia sp.)是我国重要的速生用材和绿化树种;然而,丛枝病的发生给我国的泡桐生产带来了巨大的损失。自从泡桐丛枝病病原菌被发现以来,人们对其发病机理和防治方法进行了大量研究(李秀生,1983;任国兰等, 1991;李加友,1997;范国强等,2003a;2006;何放亭等,1997;田国忠等,1994;王蕤等,1981),但由于其病原菌——植原体自身的特性,至今国内外仍不能进行体外活体培养,因此,限制了泡桐丛枝病有效防治方法的建立。众所周知,蛋白质是基因表达的最终产物,基因的信息通过mRNA转录再到合成的蛋白质。蛋白质合成抑制剂主要作用于mRNA翻译蛋白质这一过程,而核酸抑制剂主要作用于DNA的复制及mRNA的转录水平上。因此,蛋白质合成抑制剂与核酸合成抑制剂在生命代谢活动中的作用机理存在着一定差异。过去,人们用四环素和土霉素等蛋白质合成抑制剂处理泡桐(Paulownia sp.)(李秀生,1983;李加友,1997;任国兰等,1991)、木麻黄(Casuarina sp.)(容向东等,1989)、枣树(Zizyphus jujuba)(王焯等,1992)、山楂(Crataegus sp.)(张锐等,1993)、梨(Pyrus sp.)(Davies et al.,1994)和甘蔗(Sugarcane sp.) (Wongkaew et al.,2004)等患丛枝病植物后发现,这些蛋白质合成抑制剂对其症状减轻有一定作用,但绝大部分试验不能根治植物丛枝病。此外,以前研究发现,泡桐丛枝病发生与其叶片内pI 6.8、m 24 ku特异蛋白质相关(范国强等,2003a)。然而,至今国内外未见抗生素对泡桐本身蛋白质变化影响和应用核酸合成抑制剂防治泡桐丛枝病的有关研究报道。本文探讨不同浓度抗生素处理对泡桐丛枝病植原体抑制作用的差异及对丛枝病发病相关蛋白质的影响,为泡桐丛枝病防治药剂的筛选及其发病机理的阐明奠定基础。
1 材料与方法 1.1 材料及处理试验材料为河南农业大学泡桐研究所培养30 d的豫杂1号泡桐[毛泡桐×白花泡桐(Paulownia tomentosa × P. fortunei)]丛枝病组培幼苗。所用试剂为5-氟脲嘧啶(5-Fluorouracil, Fu)和利福平(Rifampicin, RF), 四环素(Tetracycline, Te)。将组培苗从基部剪断,分别接种到装有40 mL含不同浓度抗生素(表 1)的1/2MS培养基的100 mL三角瓶中,培养基中蔗糖浓度为25 g·L-1,琼脂为8 g·L-1。每瓶3个外植体,每浓度处理20瓶。在温度(25±2)℃、光强130 μmol·m-2s-1、光照时间16 h·d-1的条件下培养。30 d时,记录处理苗木的形态变化情况,并剪取适量处理幼苗叶片分别用于蛋白质双向电泳和PCR分析。
|
|
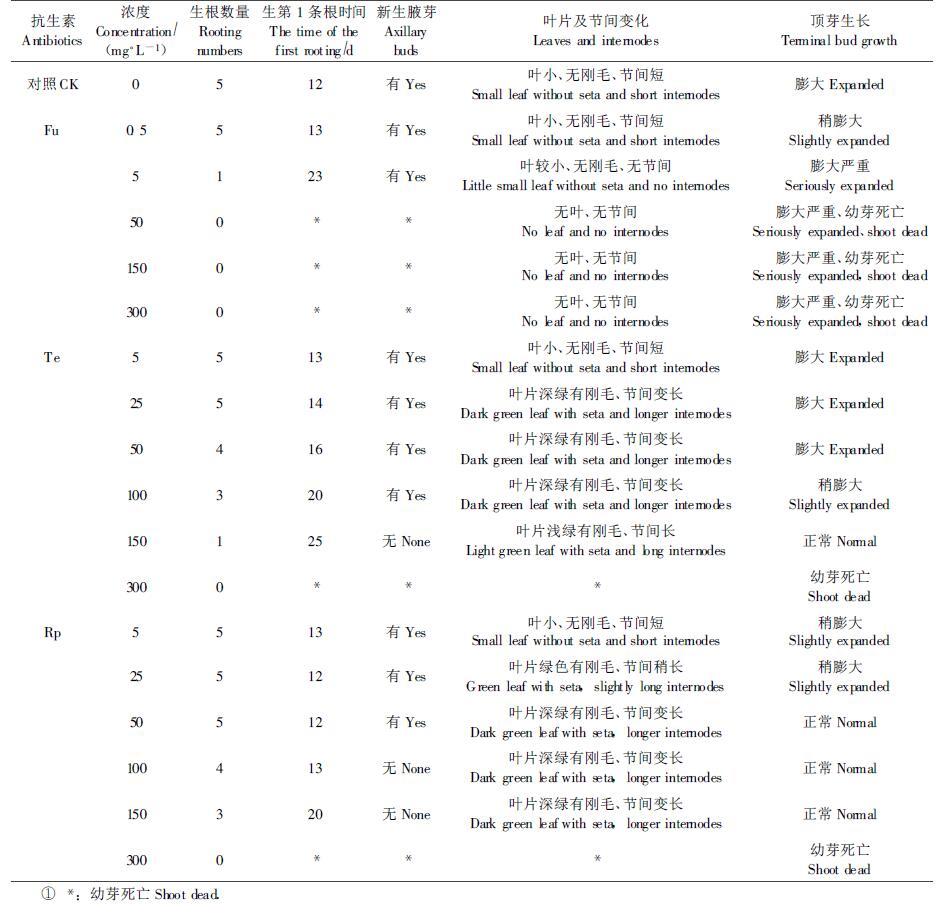
Fu和Rp及Te处理结束后,观察记录幼苗的生根时间、生根数量、幼苗新生腋芽情况、顶端膨大程度、叶片刚毛生长状况和幼苗长势等数据。
1.3 泡桐丛枝病植原体的PCR检测豫杂1号泡桐丛枝病幼苗DNA提取参照范国强等(2003b)法。PCR的基本条件是:25 μL PCR反应体系,10 × Taq酶缓冲液(10 mmol·L-1 Tris-HCl, 50 mmol·L-1 KCl, 0.1% Triton X-100, pH 9.0), 25 mmol·L-1MgCl2, 25 mmol·L-1 dNTP, 10 ng患丛枝病泡桐模板DNA,5 μmol·L-1基于植原体16S rRNA基因的16S rDNA通用引物(PrimerⅠ:5′-ACGACTGCTAAGACTGG-3′; PrimerⅡ:5′-GCGGTGTGTACAAACCCCCG-3′, 大连宝生物合成),1.25 U Taq DNA聚合酶。在PTC-200基因扩增仪上先94 ℃预变性1.5 min; 再94 ℃变性1 min, 52 ℃退火1.5 min, 72 ℃延伸2.5 min, 共30个循环;然后,72 ℃延伸10 min。PCR扩增结束后,取2 μL的PCR产物在1%琼脂糖凝胶(含0.5% EB)上40 V电泳1 h,凝胶在AlphaimagerTM 2200凝胶成像系统中观察、拍照。每一浓度抗生素处理幼苗的PCR检测试验重复3次。
1.4 泡桐叶片蛋白质的的双向电泳叶片蛋白质干粉制备及蛋白质双向电泳参照范国强等(1997;2003a)的方法。
2 结果与分析 2.1 抗生素对患丛枝病幼苗形态变化的影响1) Fu对患丛枝病苗形态的影响 由Fu对患丛枝病幼苗生长影响的结果(表 1,图 1B、C)可以看出,不同浓度Fu对豫杂一号泡桐丛枝病幼苗的形态变化有很大影响。当Fu浓度在0.5~5 mg·L-1时,随着Fu浓度的增高,患病幼苗长出第一条根所需时间逐渐变长,幼苗生根数量逐渐减少;叶片逐渐减小、节间变短,顶芽膨大越来越严重。当Fu浓度在50~300 mg·L-1时,患病幼芽没有根的形成,并且在幼芽死亡前,幼芽生长过程中顶芽和腋芽膨大严重,叶片严重退化。该结果说明,当浓度较小时,Fu对丛枝病幼苗症状的减轻作用不明显,当浓度过大时,对幼芽成活和生长造成不利影响。

|
图 1 不同抗生素处理后豫杂一号泡桐丛枝病幼苗的形态变化 Fig. 1 The morphological changes of the seedlings treated with different antibiotics A.病苗Diseased seedling(CK);B. 0.5 mg·L-1Fu;C. 100 mg·L-1Fu;D. 100 mg·L-1Te;E. 150 mg·L-1Te;F. 100 mg·L-1Rp;G. 150 mg·L-1Rp;H.健康苗Healthy seedling. |
2) Te对丛枝病幼苗形态变化的影响 不同浓度Te处理对患病幼苗形态变化影响结果(表 1,图 1D、E)表明:Te对豫杂一号泡桐丛枝病幼苗生长影响明显。当浓度小于或等于100 mg·L-1时,Te除抑制幼芽生根和推迟第1条根出现时间外,对幼苗新生腋芽和顶芽膨大等丛枝病发生形态特征没有明显的抑制作用;当浓度大于150 mg·L-1时,幼芽生根显著受到抑制,第1条根出现时间推迟明显,幼苗生长过程中没有丛生腋芽产生、顶芽不再膨大,但是幼苗新生叶片有白化现象发生;当浓度为300 mg·L-1时,Te抑制幼芽根的形成,最后导致幼芽死亡。以上结果表明:当浓度小于或等于100 mg·L-1时,Te对泡桐幼苗体内丛枝病植原体的活动起不到抑制作用; 当浓度等于或大于300 mg·L-1时,Te对幼芽产生严重的毒害作用,导致幼芽的死亡。当用150 mg·L-1Te浓度处理时,豫杂一号泡桐丛枝病幼苗体内植原体活动则可受到抑制,从而使幼苗在试验期内未出现丛枝病症状。
3) Rp对患丛枝病苗形态的影响 由Rp对豫杂一号泡桐丛枝病形态变化影响结果(表 1,图 1F、G)可以看出:Rp对患病幼苗形态变化产生了一定的作用。当Rp浓度为25~50 mg·L-1时,幼苗在第1条根出现的时间、生根数量、新生腋芽情况和节间长短等方面与患丛枝病幼苗(对照)相似,只是50 mg·L-1 Rp处理幼苗的顶芽形态上与健康幼苗相似,而25 mg·L-1 Rp处理幼苗顶芽与患病幼苗顶芽相似;当浓度为100~150 mg·L-1时,Rp抑制了幼芽生根的数量,推迟了第1条根产生的时间,同时抑制了新生腋芽的产生和顶芽的膨大,幼苗未表现丛枝病症状;当浓度为300 mg·L-1时,Rp处理的幼芽不能生根、叶片产生的褐色斑点严重,最后幼芽死亡。这些结果表明:适宜浓度的Rp对患丛枝病泡桐体内植原体的生长和繁殖有一定的抑制作用。
2.2 抗生素处理对患病幼苗体内植原体的抑制作用为了研究不同抗生素对豫杂一号泡桐丛枝病幼苗体内植原体的影响,这里只对抗生素处理后形态具有显著变化的幼苗进行了PCR扩增检测。由不同抗生素处理后的患丛枝病幼苗、健康幼苗及对照苗DNA的PCR扩增图结果(图 2)可以看出:抗生素种类和浓度对患病幼苗顶芽内植原体的抑制作用存在着一定的差异。5 mg·L-1Fu、100和150 mg·L-1Te及100 mg·L-1的Rp处理后,病苗顶芽内仍有植原体的存在,但从扩增出植原体片段量来看,150 mg·L-1Te和100 mg·L-1Rp处理后幼苗内植原体的浓度有所减少。150 mg·L-1Rp处理后,幼苗顶芽内与健康幼苗一样观察不到植原体的存在。这些结果表明:在抑制泡桐丛枝病植原体方面, Rp的作用大于Te,Te又大于Fu。结合抗生素对患病幼苗形态影响结果可知,150 mg·L-1 Te和100 mg·L-1 Rp处理病苗后,幼苗形态上虽然不呈现丛枝病症状,但是顶芽内仍有植原体存在。

|
图 2 不同抗生素处理后患病幼苗植原体16s rRNA的PCR扩增 Fig. 2 PCR of phytoplasmas 16S rRNA in the leaves with wiches' broom 1.病苗Diseased seedling; 2. 5 mg·L-1Fu; 3. 100 mg·L-1Te; 4. 150 mg·L-1Te; 5. 100 mg·L-1Rp; 6. 150 mg·L-1Rp; 7.健康苗Healthy plantlets; 8.DNA分子量标准DNA maker. |
抗生素处理后,病苗叶片蛋白质发生了较为明显的变化(图 3)。为了阐明抗生素处理对泡桐丛枝病发生相关蛋白质(范国强等,2003)的影响,这里只对抗生素处理后丛枝病幼苗形态变化明显的幼苗叶片进行蛋白质双向电泳分析。用Te处理后,即使形态呈现无丛枝病症状的豫杂一号泡桐患病幼苗叶片蛋白质双向电泳图谱中也观察不到pI 6.8,m 24 ku蛋白质。在Rp处理的无丛枝病症状幼苗叶片蛋白质图谱中,100 mg·L-1Rp处理幼苗叶片内也没有pI 6.8,m 24 ku的存在,而150 mg·L-1 Rp处理幼苗叶片内则出现该蛋白质。此外,未处理患病幼苗叶片内也没有该蛋白质的存在,而在健康幼苗叶片内存在该蛋白质。这表明150 mg·L-1Rp处理患病苗可诱导体内pI6.8,m24 ku蛋白质的重现。因而推测,150 mg·L-1 Rp处理可能清除患病幼苗内存在的植原体,使由于植原体干扰而导致降低或停滞合成该蛋白质的基因重新激活,幼苗不呈现丛枝病症状。

|
图 3 抗生素处理后病苗叶片蛋白质的双向电泳图谱 Fig. 3 2-dimensional electrophoresis of proteins related to witches' broom in the treated plantlets with antibiotics A.病苗Diseased seedling;B. 150 mg·L-1Te;C. 150 mg·L-1Rp;D.健康苗Healthy seedling. |
植原体是一类专性寄生于植物细胞内的无细胞壁、至目前仍不能进行离体培养的微生物(Agrios, 1997)。据统计,植原体可导致世界范围内600余种植物的生长发育过程发生异常,出现不同的丛枝病症状(Nishigawa et al.,2002),给农业、林业和花卉种植造成了巨大的经济损失。以前,人们试图用抗生素杀死寄主体内存在的植原体(李加友,1997;李秀生,1983;任国兰等,1994;容向东等,1989;张锐等,1993;Davies et al.,1994),但效果大部分都不理想。本试验通过不同浓度Fu和Rp对豫杂一号泡桐丛枝病幼苗形态等方面影响的研究发现,所有浓度Fu处理后,病苗丛枝病症状不能消失,而用150 mg·L-1Rp处理后,病苗症状消失,并且用PCR扩增也检测不到植原体的存在。该结果可能与Rp对原核生物核酸合成的抑制作用比Fu更强有关,适宜浓度的Rp可清除豫杂一号泡桐丛枝病幼苗内的植原体。此外,本结果还表明:与泡桐丛枝病发生特异相关的pI 6.8,m 24 ku蛋白质(范国强等,2003a)在患丛枝病豫杂一号泡桐幼苗及用3种抗生素处理后仍有植原体存在的幼苗内观察不到,而在150 mg·L-1Rp处理后的幼苗中又重新出新。该结果一方面再次说明该蛋白质与泡桐丛枝病发生密切相关,另一方面也表明植原体侵入可能导致该蛋白质对应基因的沉默。
在四环素防治泡桐丛枝病方面,目前的研究结果存在一定的差异(李加友,1997;李秀生,1983;任国兰等,1991),其原因可能与不同研究者所用泡桐种类和试验条件不同有一定的关系。本试验中,100 mg·L-1Te处理豫杂一号泡桐丛枝病幼苗的丛枝病症状不能消除, 150 mg·L-1Te处理的幼苗丛枝病症状消失,但幼苗体内仍有植原体的存在。该结果的出现可能与Te主要作用于mRNA到蛋白质的翻译过程的生理生化反应密切相关(Turner et al., 1997)。也就是说,低浓度Te抑制不了患病幼苗体内植原体通过蛋白合成产生的干扰作用;适宜浓度的Te,虽然在一定程度上抑制了植原体的繁殖及作用,使患丛枝病幼苗暂时不呈现丛枝病症状,但植原体仍存在于寄主内,一旦Te药效丧失,随时间延长幼苗体内植原体浓度逐渐增大,最后丛枝病症状重新出现;过高浓度的Te则在抑制植原体蛋白质合成的同时,也抑制了泡桐生理生化代谢所需蛋白质的合成,影响泡桐的正常生长。类似现象在患丛枝病月见草属植物中出现(Sears et al.,1989)。因此,Te杀死植原体的结果还有待于进一步证实。
范国强, 冯志敏, 翟晓巧, 等. 2006. 植物生长调节物质对泡桐丛枝病株幼苗形态和叶片蛋白质含量变化的影响. 河南农业大学学报, 40(2): 137-141. DOI:10.3969/j.issn.1000-2340.2006.02.007 |
范国强, 蒋建平. 1997. 泡桐叶片蛋白质提取方法的研究. 植物学通报, 14(3): 61-64. |
范国强, 李有, 郑建伟, 等. 2003a. 泡桐丛枝病发生相关蛋白质的电泳分析. 林业科学, 39(2): 119-122. |
范国强, 马新业. 2003b. 泡桐叶片DNA提取. 河南科学, 21(2): 172-175. |
何放亭, 戴群, 武红巾, 等. 1997. C/A值与甘薯丛枝病症状发生的关系. 植物病理学报, 27(1): 43-46. DOI:10.3321/j.issn:0412-0914.1997.01.012 |
李加友. 1997. 抗生素对组培条件下植物枝原体作用的初探. 南京农专学报, 13(4): 25-31. |
李秀生. 1983. 泡桐丛枝病防治技术的研究. 河南农学院学报, 17(2): 49-58. |
任国兰, 李秀生, 郑兰长, 等. 1991. 新农药"去丛灵"的应用效果研究. 河南农业大学学报, 25(1): 51-55. |
容向东, 张景宁. 1989. 木麻黄黄化丛枝病的研究. 植物病理学报, 19(4): 217-221. |
田国忠, 黄钦才, 袁巧平, 等. 1994. 感染MLO泡桐组培苗代谢变化与致病机理的关系. 中国科学:B, 24(5): 484-490. |
王蕤, 孙秀琴, 王守宗, 等. 1981. 激素对泡桐丛枝病发生的影响. 林业科学, 17(3): 281-286. |
王焯, 周佩珍, 郭裕新, 等. 1992. 四环素消毒枣疯病接穗研究. 植物保护学报, 19(1): 62. |
张锐, 王敏, 金开璇, 等. 1993. 土霉素对山楂丛枝病的治疗试验. 北京农业科学, 6(11): 29-31. |
Agrios G N. 1997. Plant diseases caused by Mollicutes: phytoplasmas and spiroplasmas//Agrios G N. Plant Pathology(4th Edition). New York: Academic Press
|
Davies D L, Clark M F. 1994. Maintenance of mycoplasma-like organisms occurring in Pyrus species by micropropagation and their elimination by tetracycline therapy. Plant pathology, 43: 819-823. DOI:10.1111/j.1365-3059.1994.tb01626.x |
Nishigawa H, Oshima K, Kakizawa S, et al. 2002. A plasmid from a non-insect-transmissible line of a phytoplama lacks two open reading frames that exist in the plasmid from the wild-type line. Gene, 298: 195-201. DOI:10.1016/S0378-1119(02)00975-7 |
Sears B B, Klomparens K L. 1989. Leaf tip cultures of the evening primrose allow stable, aseptic culture of mycoplasmas——like organism. Canada Journal of Plant Pathology, 11: 343-348. DOI:10.1080/07060668909501077 |
Turner P C, MeLennan A G, Bates A D, et al. 1997. Instant Notes in Molecular Biology. United Kingdom: Scientific Publishers Limited, 247-261.
|
Wongkaew P, Fletcher J. 2004. Sugarcane white leaf phytoplasma in tissue culture: long-term maintenance, transmission, and oxytetracycline remission. Plant Cell Reports, 23: 426 434. |