 2005, Vol. 41
2005, Vol. 41文章信息
- 曾德慧, 陈广生, 陈伏生, 赵琼, 冀小燕.
- Zeng Dehui, Chen Guangsheng, Chen Fusheng, Zhao Qiong, Ji Xiaoyan.
- 不同林龄樟子松叶片养分含量及其再吸收效率
- Foliar Nutrients and Their Resorption Efficiencies in Four Pinus sylvestris var. mongolica Plantations of Different Ages on Sandy Soil
- 林业科学, 2005, 41(5): 21-27.
- Scientia Silvae Sinicae, 2005, 41(5): 21-27.
-
文章历史
- 收稿日期:2004-07-07
-
作者相关文章


养分再吸收(nutrient resorption)是指养分从衰老叶片中转移并被运输到植物其他组织的过程(Killingbeck, 1986; Aerts, 1996)。养分从衰老的叶片或其他树木组织中的再迁移、再吸收或再分配使得养分在植物体内的存留时间延长,从而可以提供树木新的生物量生产所需的大部分养分(Sollins et al., 1980; Meier et al., 1985)。养分的再吸收不仅使树木很少受到土壤养分可利用性波动的影响,而且使得凋落物分解时的养分淋溶量减少,从而减缓养分从整个系统的损失。因此,养分再吸收不仅是植物对养分贫瘠环境的一种适应进化机制(Aerts, 1990; May et al., 1992; Santa et al., 2001),也是植物保存养分、增强竞争力、提高养分吸收能力和生产力的重要策略之一。这个过程控制了养分从衰老植物组织中移动和回收,以及这些养分向多年生的储存器官的运输(Chapin, 1980; Killingbeck, 1986)。Berendse等(1987)和Aerts(1997)认为养分贫瘠生境的植物的共同特征表现为具有高的养分保存效率,而并不是因为具有很强的对土壤养分的吸收能力。养分再吸收大小通常以养分再吸收效率表示,它是指叶片衰老过程中再吸收的养分量占叶片衰老前的养分量的百分率(Killingbeck, 1986),不同物种的养分再吸收效率差异很大,5%~80%的植物叶片N再吸收效率和0~95%的叶片P再吸收效率都有报道(Aerts et al., 2000)。
20世纪30年代,科学家们就已经注意到叶片养分在衰老过程中存在着再吸收的现象(Killingbeck, 1986),到目前仍有大量养分再吸收的研究报导(Lal et al., 2001; van Heerwaarden et al., 2003a)。但是,这些研究大都集中在对不同物种的叶片养分再吸收效率的比较上,对于同一物种不同年龄阶段的养分再吸收效率的研究非常少(Helmisaari, 1992a)。国内对植物养分再吸收效率的研究始于20世纪90年代初,除一些学者介绍国外研究进展外(苏波等,2000; 邢雪荣等,2000),研究工作主要集中在杨树(沈善敏等,1992; 1993;陈欣等,1995; 宇万太等,1995; 徐福余等,1997)和沙地柏(Sabina vulgaris)(何维明等,2002)等少数树种,而且都着重于植物叶片养分再吸收效率的现状研究,对植物养分再吸收效率差异的机制和生态学意义缺乏进一步的分析和探讨。
20世纪50年代中期,樟子松(Pinus sylvestris var. mongolica)引种到科尔沁沙地东南缘章古台地区后,表现了较强的适应性(王康富等,1958),早期生长速度明显高于原产地,而且也比油松(P. tabulaeformis)和赤松(P. densiflora)高,因此,樟子松很快引种推广到我国北方许多沙地,成为我国北方干旱半干旱地区的一个主要造林树种(曾德慧等, 1997)。但是,20世纪80年代末章古台地区的樟子松林出现了一定面积的衰退甚至死亡的现象(曾德慧等, 1996)。姜凤岐等(1996;1997)根据沙地樟子松人工林的生长发育规律,推测章古台地区的樟子松在46~56年生进入成熟状态,因此,章古台樟子松的早衰和死亡,引起了人们的极大关注。关于樟子松的衰退原因存在很多推测(焦树仁, 2001),由于缺乏试验数据,对樟子松衰退机制的研究一直未能取得理想的进展。本项研究主要从养分角度来探讨樟子松的衰退问题,目的是通过叶片养分再吸收效率来比较不同年龄樟子松对养分的保存能力,检验樟子松对贫瘠养分生境的适应能力是否是随年龄的增加而降低,为该地区樟子松林营林措施提供理论依据。
1 材料与方法 1.1 试验地概况试验地位于辽宁省彰武县章古台(42°43′N,122°22′E),科尔沁沙地的东南缘,平均海拔226.5 m,地形为低缓沙丘。本区属于亚湿润干旱区,年均温5.9 ℃,≥10 ℃的年积温2 890 ℃,降水量496 mm,蒸发量1 700 mm左右,相对湿度58%,干燥度3.643,极端最低温-29.5 ℃,极端最高温37.2 ℃,无霜期约154 d,地下水位3.4~6.8 m。土壤类型主要为生草沙土和流动风沙土,养分含量非常低(焦树仁, 2001)。
章古台地区是草原和森林分布交错区,草本植物多样性比较丰富,而乔木树种相对较少。乔木树种(大部分为引种)主要有:樟子松、油松、赤松、红皮云杉(Picea koraiensis)、侧柏(Platycladus orientalis)、彰武小钻杨(Populus xiaozhuanica)、新疆杨(P. alba var. pyramidalis)、旱柳(Salix matsudana)、蒙古栎(Quercus mongolica)、元宝槭(Acer truncatum)、白榆(Ulmus pumila)、刺槐(Robinia pseudoacacia)等。
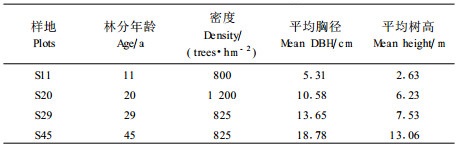
在地形条件相似、相互间距小于3 km的11、20、29、45年生樟子松纯林中,各选取1块20 m×20 m、地势较平坦、树木生长状况良好的林地作为试验样地,所选试验样地距离林分边缘均大于50 m。于2003年8月对样地内树木进行每木检尺,获得林分的基本特征参数(表 1),表 1中S11、S20、S29、S45分别表示11、20、29、45年生的樟子松样地。
|
|
本次试验樟子松针叶样品第1次的采集时间是2002年10月27日,第2次于2003年4月15日进行,以后每月中旬取样1次,止于2003年10月15日,取样8次。根据每木检尺资料,在所设样地中找出接近平均胸径和树高、生长良好的树木作为样树,进行标记。每次取样的树木为15株,3株邻近样树上所取的针叶组成1个重复。考虑到树木的叶片养分含量在不同高度有差异,本次研究选取树冠中部的叶片,以代表整个树冠层的叶片。每株样树东、南、西、北方向的一级枝条各取3个小枝(直接与树干相连的枝条定义为一级枝条,直接与一级枝条相连的为二级枝条,而直接与二级枝条相连的为三级枝条),以消除叶片养分含量因枝条方向的不同而产生的取样误差(Helmisaari, 1992a; 1992b)。每次取样时,随机于3株邻近样树选定的一级枝上用高枝剪剪下3条三级枝条,把4个方向剪下的三级枝条混合,然后按每个枝条的各龄级针叶分开摘取,这样每3株样树具有各年龄级的针叶样品各1个,重复数为5。将样品带至实验室后,于每个龄级的针叶中按各龄级针叶在三级枝条上的数量比例关系混合各龄级针叶,组成混合样品,这样每次取样每一样地就同时具有5个混合样品。考虑到多次对同一一级枝条上的三级枝条取样会影响取样枝以及枝条的其他三级枝条叶片的养分含量,同时也会影响该株树木存留的其他枝条上的针叶养分含量,因此,在试验过程中对样树的一级枝条重新进行了3次选择,样树也重新选择了1次,鉴于选择样树是随机的,样树的重新选择对测量结果不会带来很大误差。整个研究过程中每个样地取样树木为30株。
1.2.2 针叶季节生长的测定理论上,测量针叶在一年中的生长时,跟踪观测固定标记的针叶是最准确的方法(Van Heerwaarden et al., 2003b),这种方法一方面可以不破坏树木,另一个重要方面可以准确测定生长量,但是,由于实际操作很难,实际测量时采取的是测定固定标记的一级枝条上的针叶的生长量。在每个样地选择的15株样树中再选取3株,每株各标记东南西北方向的3个枝条(总共12个枝条),并于2003年5、7、9和10月进行了4次取样。每个枝条取各龄级针叶10束(2针1束)带回实验室,针叶烘干(70 ℃,48 h)后,用千分尺测定针叶长度和宽度(针叶中间部分),并测定单根针叶平均干质量,各龄级针叶各生长指标的平均值即为混合样品的针叶各生长指标的平均值。根据焦树仁(1989)模拟的樟子松针叶叶面积经验公式S=2.57R(L-0.116 7)求算叶面积,式中:S为叶面积(cm2),R为针叶宽度(cm),L为针叶长度(cm)。
1.2.3 凋落物收集2002年10月底在每一块样地中随机放置5个长、宽、高都为50 cm的凋落物收集木箱,这种尺寸的木箱足以防止凋落物从箱内飘出或地上凋落物飘进。木箱底部具有微小间隙以供雨水迅速渗漏,木箱内部底部及其周边铺盖尼龙网袋以固定凋落物并使凋落物不能接触到木箱底部可能具有的生物,从而抑制凋落物在每个月的收集间隔期的分解。2003年4月中旬第1次收集凋落物量,以后每月收集1次,直至2003年10月底。每次凋落物取回后,置于70 ℃的烘箱中烘干48 h,然后立即对凋落物进行分类,分别测定针叶、枯枝、花粉、树皮质量,同时,取100束凋落针叶测定凋落叶片平均单叶质量,对凋落量最大的9月和10月的凋落叶片烘干后混合随机取出100束针叶进行单叶平均长度和宽度测定。测定完这些指标后,对凋落针叶进行粉碎,过0.5 mm筛,针叶粉末装入密封袋中,标记后保存留待测定。
1.3 样品养分含量测定所有植物样品在测定养分含量前,都要再次烘干8 h(70 ℃),冷却后立即称样,然后进行其他操作。常量元素含量采用常规方法测定。叶片N含量采用浓H2SO4-H2O2消煮、凯氏法测定,消煮液同时用以P和K含量的测定; 叶片P含量采用酸溶-钼锑抗比色法测定; 叶片K含量采用火焰光度法测定; 植物有机碳的测定采用油浴-K2Cr2O7容量法; 针叶样品在550 ℃下灰化至灰白色,然后,以HCl浸提,采用EDTA容量法测定Ca和Mg含量。
1.4 叶片平均养分含量及养分再吸收效率的计算本次研究中对针叶进行了8次取样,用2003年的7次取样样品(从2003年4月至10月)测定的叶片养分含量计算2003年樟子松绿色叶片(凋落前的叶片)的平均养分含量。
由于常绿树种在其生活史中多个年龄叶片并存,因此,叶片凋落前的养分含量难以确定,有的研究者以常绿树种一年中生长季结束的8、9或10月的叶片养分为凋落前的叶片养分(Helmisaari, 1992a; 1992b),但是,在大多数研究中都把凋落前的并存叶片(各年龄叶片混合)的最高养分含量作为常绿树种凋落前的叶片养分含量(May et al., 1992; Wright et al., 2003)。这种方法计算的养分再吸收效率被认为是该树种的年最大养分再吸收效率,本次研究也采用这种方法,因此,获得的养分再吸收效率也是一种最大养分再吸收效率值。根据章古台地区樟子松针叶凋落的季节性特征,选择针叶凋落量最大的9、10月的叶片凋落物混合后,测定养分含量,即叶片凋落后的平均养分含量。
养分是构成植物体质量的一部分,因此,衰老过程中叶片质量会发生变化。同时,叶片衰老过程中,叶片逐渐干枯,会使叶片产生收缩,阔叶树种叶片的收缩更为明显。因此,为了消除叶片衰老过程中的质量下降和叶面积收缩产生的养分再吸收效率计算误差,出现了多种养分再吸收效率的表述方法:基于单叶质量的叶片养分再吸收效率(Rmass)、基于单位叶面积的再吸收效率(Rarea)以及基于整个树冠的叶片养分再吸收效率(Rcanopy)。上述3种养分再吸收效率的计算方法分别为:
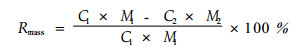
|
(1) |
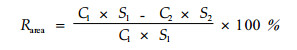
|
(2) |
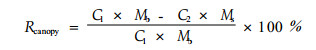
|
(3) |
式中:下标1、2分别表示凋落前的叶片和凋落时的该叶片; C为单位叶片质量的养分含量(%); S为单位叶面积的质量(比叶质量specific leaf mass)(g·cm-2); M1、M2分别为凋落前与凋落后的该叶片的单叶质量(g); Mb是凋落前的整个树冠的叶片总质量(kg·hm-2),Ms是凋落的叶片总质量(kg·hm-2)。
由于研究树种主要为常绿树种,凋落的叶片只是树木整个树冠叶片的一部分,因此,公式(3)不适于本次研究。本项研究主要以公式(1)和(2)来表述并比较养分再吸收效率的大小,文中涉及叶片质量的各种计算都是基于干质量。
2 结果与分析 2.1 凋落前后树木叶片特征变化表 2列出了樟子松叶片脱落前后的单叶质量、单叶面积、比叶质量、碳含量的变化。从表中可见,不同林龄的樟子松叶片脱落后单叶质量下降了4%~10%,下降幅度随林龄增加而逐渐缩小,产生这种差异的原因可能是年龄相对较小的林木,生长快具有较高的光合速率,因而,可溶性蛋白质含量较高,在落叶前蛋白质和其他细胞组分的水溶性增强,易于从叶片中转移,导致叶片质量下降很多(Aerts, 1997)。叶片凋落后,单叶面积减少了4.8%~5.3%,不同树龄之间变化差异不大; 比叶质量变化为-5.3%~1.1%,减少幅度随树龄增加逐渐下降,到45年生时凋落叶片的比叶质量比凋落前稍有增加。叶片凋落后,不同年龄的树木叶片中的碳含量都有所增加,可能是由于叶片中其他养分发生再吸收后,叶片中碳的比例增大了。
|
|
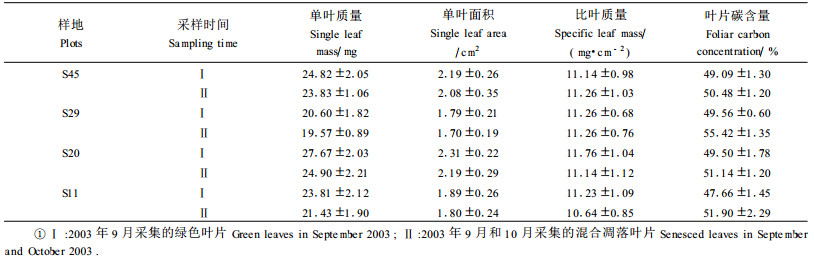
对7次(从2003年4月到10月每月1次)取的绿色针叶的养分含量测定后,取平均值,即为叶片凋落前(绿色叶片)的平均养分含量(表 3)。从表中可见20年生的樟子松针叶的平均N含量最高(15.37 g·kg-1),而11年生的则最低(12.84 g·kg-1)。整体上樟子松叶片的N含量并未表现出随树木年龄变化而出现有规律的增减。造成这种现象的可能原因是叶片的养分含量受到其生长立地土壤养分可利用状况和土壤含水量等的差异影响。20年生的樟子松针叶P含量显著高于29和45年生的树木(P<0.05),而与11年生的树木差异不明显(P>0.05)。4个年龄樟子松的针叶K含量之间没有明显差异,反映了年龄的变化没有改变樟子松叶片对K的吸收和保存生理特征。11年生樟子松叶片Ca含量显著大于其他3个年龄樟子松。29和20年生樟子松叶片Ca含量最低,而Mg含量最高。樟子松树木叶片的N/P表现出随年龄增加而增加的趋势,反映了随年龄增加樟子松叶片的N/P出现了一种调整,这可能是樟子松为适应章古台土壤可利用N、P供应变化的一个重要改变。
|
|
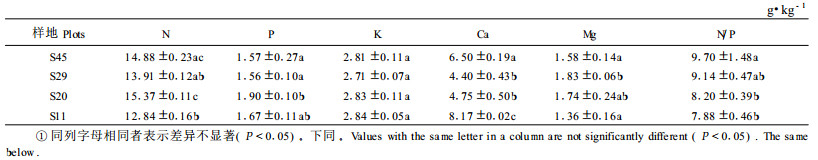
表 4中列出了4个林龄樟子松在9月和10月的凋落叶片的平均养分含量,从中可以发现11年生樟子松叶片凋落物中N含量显著低于其他3个年龄的树木(P<0.05),而45年生、29年生和20年生樟子松叶片凋落物中的N含量没有表现出显著性差异(P>0.05)。45年生樟子松凋落叶片中的P含量显著大于其他3个年龄树木(P<0.05),而这3个年龄樟子松树木之间没有显著差别(P>0.05)。11年生和20年生樟子松叶片凋落物中的K含量显著低于45年生和29年生的树木(P<0.05),而29年生樟子松凋落叶片中的K含量显著低于45年生樟子松(P<0.05),表现出了凋落叶片中的K含量随着年龄增加而逐渐增加的趋势。而凋落叶片的Ca含量,则与K含量趋势正好相反,表现出了随着年龄增加而逐渐降低的趋势。11年生樟子松凋落叶片中的Mg含量显著低于其他3个年龄,而这3个年龄间的差异并不显著(P>0.05)。
|
|
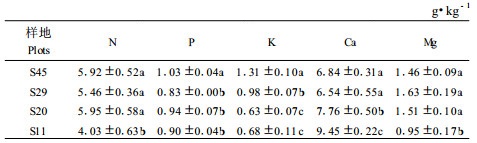
从图 1A、B可以看出基于单位叶面积和单叶质量的樟子松叶片养分再吸收效率表现出相同的规律。因此,下面将以基于单叶质量的叶片养分再吸收效率(图 1B)进行分析。45、29、20年生樟子松树木叶片的N再吸收效率差异不明显,其再吸收效率都在65%左右,但这3个年龄树木的再吸收效率都显著低于11年生的樟子松(73%); 45年生樟子松叶片P的再吸收效率显著低于其他3个年龄的树木,叶片P的再吸收效率表现为:20年生>11年生>29年生>45年生樟子松。11和20年生樟子松树木叶片K的再吸收效率(分别为79%和82%)显著高于29和45年生树木(分别为68%和62%),表现出了随年龄增加叶片K养分再吸收效率逐渐降低的趋势; 4个年龄樟子松叶片凋落后Ca含量都表现出了不同程度的增加,养分再吸收效率为负值,45和11年生樟子松树木叶片对Ca的保存能力显著强于20和29年生的樟子松,45年生樟子松单叶质量中Ca累积率仅为1%,而29和20年生的樟子松累积率分别为41%和47%;4个年龄的樟子松叶片凋落后单叶质量的Mg含量有所下降,并且表现出了随年龄增加再吸收效率逐渐降低的趋势。

|
图 1 基于单位叶面积(A)和基于单叶质量(B)的不同年龄樟子松叶片的养分再吸收效率 Fig. 1 Foliar nutrient resorption efficiency based on leaf area (A) and on single leaf mass (B) of P. sylvestris var. mongolica trees of different ages |
从总体上看,45年生樟子松树木叶片对易于迁移的N、P、K、Mg元素的再吸收效率都显著低于其他3个年龄,而11、20年生樟子松表现了最大的再吸收效率,因此,总体上樟子松树木叶片N、P、K、Mg的再吸收效率表现了随年龄增加而减少的趋势。4个年龄的樟子松叶片对N、P、K、Ca、Mg的平均再吸收效率分别为67%、57%、72%、-23%、21%,树木叶片对5种元素的再吸收效率表现为K>N>P>Mg>Ca。
3 结论与讨论养分再吸收对于营养元素的循环具有重要意义,许多研究(Grime, 1979; Chapin, 1980; Berendse et al., 1987; Aerts, 1990; May et al., 1992)表明,在养分含量低的生境中,低的养分损失率能够使植物种群保持健康生长。在叶片衰老过程中再吸收的养分可以直接供应植物的继续生长,从而减少植物对土壤养分吸收的依赖性(Aerts, 1996)。凋落叶片中那部分未发生再吸收的养分,则主要通过凋落物的分解回收到土壤中供植物吸收,而重新被植物利用。由于针叶树种凋落物的养分释放过程往往需要几年的时间(Staaf et al., 1982; Berg, 1986),在短期内凋落物中的养分无法被植物所吸收。因此,养分再吸收效率能够表述植物叶片的相对养分保存效率(Killingbeck, 1986; Aerts, 1996)。章古台地区的人工林生态系统长期受到强烈人为干扰(放牧、凋落物收集)的影响,林内每年的树木凋落物的70%~90%都被人为收集走了,同时,林内的草本植物,也因长期的放牧活动,生物量极低(焦树仁, 1989)。每年都有大量的养分因为这2种原因以及林木生物量的收获而从生态系统中流失,在养分贫瘠的沙地养分不断流失而又得不到补充的情况下,系统的养分供应迟早会成为限制植物生长的主要因素,因此,这一生态系统中的植物叶片的养分再吸收效率就将直接影响到整个系统的养分水平、植物生产力水平、林分和土壤的衰退速度及系统的健康状况。
樟子松叶片的N、P、K、Mg再吸收效率表现出了随年龄增加而降低的趋势,表明随着樟子松年龄的增加,樟子松树木的N、P、K、Mg养分保存能力逐渐降低,而这种养分保存能力的降低表明了随着年龄的增加樟子松对章古台生境的适应性逐渐降低,樟子松在对养分的保存上表现出了一种衰退特征。为此,建议在章古台地区对樟子松人工林采取禁牧和封育的经营措施,防止凋落物的大量损失而加剧养分的亏缺; 另一方面,对衰退樟子松及时进行更新,同时着手营造混交林,以维护生态系统的稳定性和持续性。
陈欣, 宇万太, 张璐, 等. 1995. 不同施肥杨树主要营养元素内外循环比较研究Ⅱ.施肥对落叶前后杨树叶片营养元素浓度及贮量的影响. 应用生态学报, 6(4): 346-348. |
何维明, 张新时. 2002. 沙地柏对毛乌素沙地3种生境中养分资源的反应. 林业科学, 38(5): 1-6. DOI:10.3321/j.issn:1001-7488.2002.05.001 |
姜凤岐, 曾德慧, 范志平, 等. 1996. 沙地樟子松林单木生长的研究. 应用生态学报, 7(增刊): 1-5. |
姜凤岐, 曾德慧, 朱教君. 1997. 固沙林的经营基础与技术对策. 中国沙漠, 17(3): 250-254. |
焦树仁. 1989. 章古台固沙林生态系统的结构和功能. 沈阳: 辽宁科学技术出版社.
|
焦树仁. 2001. 辽宁章古台樟子松固沙林提早衰退的原因与防治措施. 林业科学, 37(2): 131-138. |
沈善敏, 宇万太, 张璐, 等. 1992. 杨树主要营养元素内循环及外循环研究Ⅰ.落叶前后各部位养分浓度及养分贮量变化. 应用生态学报, 3(4): 296-301. |
沈善敏, 宇万太, 张璐, 等. 1993. 杨树主要营养元素内循环及外循环研究Ⅱ.落叶前后养分在植株体内外的迁移和循环. 应用生态学报, 4(1): 27-31. |
苏波, 韩兴国, 黄建辉, 等. 2000. 植物的养分利用效率(NUE)及植物对养分胁迫环境的适应策略. 生态学报, 20(2): 335-343. DOI:10.3321/j.issn:1000-0933.2000.02.026 |
王康富, 赵兴梁, 杨喜林. 1958. 东北西部沙荒地区引种松树的三年试验. 林业科学, (1): 1-21. |
邢雪荣, 韩兴国, 陈灵芝. 2000. 养分利用效率研究综述. 应用生态学报, 11(5): 785-790. DOI:10.3321/j.issn:1001-9332.2000.05.033 |
徐福余, 王力华, 李培芝, 等. 1997. 若干北方落叶树木叶片养分的内外迁移Ⅰ.浓度和含量的变化. 应用生态学报, 8(1): 1-6. |
宇万太, 陈欣, 张璐, 等. 1995. 不同施肥杨树主要营养元素内外循环比较研究Ⅰ.施肥对杨树生物量及落叶前后N内外循环的影响. 应用生态学报, 6(4): 341-345. |
曾德慧, 姜凤岐, 范志平, 等. 1996. 沙地樟子松人工林稳定性的研究. 应用生态学报, 7(4): 337-343. |
曾德慧, 姜凤岐, 范志平, 等. 1997. 樟子松沙地人工林直径分布模拟. 应用生态学报, 8(3): 231-234. DOI:10.3321/j.issn:1001-9332.1997.03.002 |
Aerts R, Chapin F S Ⅲ. 2000. The mineral nutrition of wild plants revisited: a re_evaluation of processes and patterns. Adv Ecol Res, 30: 1-67. |
Aerts R. 1990. Nutrient use efficiency in evergreen and deciduous species from heathlands. Oecologia, 84: 391-397. DOI:10.1007/BF00329765 |
Aerts R. 1996. Nutrient resorption from senescing leaves of perennials: are there general patterns?. J Ecol, 84: 597-608. DOI:10.2307/2261481 |
Aerts R.. 1997. Nitrogen partitioning between resorption and decomposition pathways: a trade-off between nitrogen use efficiency and litter decomposability?. Oikos, 80: 603-606. DOI:10.2307/3546636 |
Berendse F, Aerts R. 1987. Nitrogen-use-efficiency: a biologically meaningful definition?. Funct Ecol, 1: 293-296. |
Berg B. 1986. Nutrient release from litter and humus in coniferous forest soils —a mini review. Scand J For Res, 1: 359-369. DOI:10.1080/02827588609382428 |
Chapin F S III. 1980. The mineral nutrition of wild plants. Annu Rev Ecol Syst, 11: 233-260. DOI:10.1146/annurev.es.11.110180.001313 |
Grime J P. 1979. Competition and the struggle for existence. In: Anderson R M, Turner B D, Taylor L R, (eds). Population dynamics. Symposia of the British Ecological Society 20. Oxford: Blackwell Scientific Publications, 123-139
|
Helmisaari H S. 1992a. Nutrient retranslocation in three Pinus sylvestris stands. For Ecol Manag, 51: 347-367. DOI:10.1016/0378-1127(92)90334-6 |
Helmisaari H S. 1992b. Nutrient retranslocation within the foliage of Pinus sylvestris. Tree Physiol, 10: 45-58. DOI:10.1093/treephys/10.1.45 |
Killingbeck K T. 1986. The terminological jungle revisited: making a case for use of the term resorption. Oikos, 46: 263-264. DOI:10.2307/3565477 |
Lal C B, Annapurna C, Raghubanshi A S, et al. 2001. Effect of leaf habit and soil type on nutrient resorption and conservation in woody species of a dry tropical environment. Can J Bot, 79: 1066-1075. |
May J D, Killingbeck K T. 1992. Effects of presenting nutrient resorption on plant fitness and foliar nutrient dynamics. Ecology, 73: 1868-1878. DOI:10.2307/1940038 |
Meier C E, Grier C C, Cole D W. 1985. Below and aboveground N and P use by Abies amabilis stands. Ecology, 66: 1928-1942. DOI:10.2307/2937389 |
Santa Regina I, Leonardi S, Rapp M. 2001. Foliar nutrient dynamics and nutrient-use efficiency in Castanea sativa coppice stands of southern Europe. Forestry, 74: 1-10. DOI:10.1093/forestry/74.1.1 |
Sollins P, Grier C C, McCorison F M, et al. 1980. The internal element cycles of an old-growth Douglas-fir ecosystem in western Oregon. Ecol Monogr, 50: 261-285. DOI:10.2307/2937252 |
Staaf H, Berg B. 1982. Accumulation and release of plant nutrients in decomposing Scots pine needle litter Ⅱ. Long-term decomposition in a Scots pine forest. Can J Bot, 60: 1560-1568. |
Van Heerwaarden L M, Toet S, Aerts R. 2003a. Nitrogen and phosphorus resorption efficiency and proficiency in six subarctic bog species after 4 years of nitrogen fertilization. J Ecol, 91: 1060-1070. DOI:10.1046/j.1365-2745.2003.00828.x |
Van Heerwaarden L M, Toet S, Aerts R. 2003b. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions. Oikos, 101: 664-669. DOI:10.1034/j.1600-0706.2003.12351.x |
Wright I J, Westoby M. 2003. Nutrient concentration, resorption and lifespan: leaf traits of Australian sclerophyll species. Funct Ecol, 17: 10-19. DOI:10.1046/j.1365-2435.2003.00694.x |