 2005, Vol. 41
2005, Vol. 41文章信息
- 巩合德, 王开运, 杨万勤.
- Gong Hede, Wang Kaiyun, Yang Wanqin.
- 川西亚高山3种森林群落穿透雨和茎流养分特征研究
- Nutrient Characteristics of Throughfall and Stemflow in Three Forests at the Subalpine of Western Sichuan
- 林业科学, 2005, 41(5): 14-20.
- Scientia Silvae Sinicae, 2005, 41(5): 14-20.
-
文章历史
- 收稿日期:2003-12-30
-
作者相关文章


2. 华东师范大学城市化过程和生态恢复重点实验室 上海 200062;
3. 四川农业大学林学院 雅安 625014
2. Urbanization Process and Ecological Restoration Key Laboratory, East China Normal University Shanghai 200062;
3. Faculty of Forestry Sichuan Agricultural University Ya'an 625014
大气降雨是森林生态系统养分输入的主要形式之一,甚至是养分缺乏地区生态系统养分输入的全部形式。因此,降雨输入的营养元素不仅对于森林群落的生长和演替具有重要意义,而且是森林生态系统生物化学循环的重要组成部分(Rodrigo et al., 2003)。由于森林水文过程中的养分动态对于研究系统的养分输入、输出以及评价养分循环是必不可少的(田大伦等,2002),因此有关各种森林生态系统大气降雨、穿透雨和树干茎流的养分动态和输入受到了国内外学者的普遍关注(Lindberg et al., 1985; 樊后保,2000; 黄建辉等,2000; Aboal et al., 2002; Fan et al., 2001, 陈永瑞等,2003)。但由于林冠截获的干沉降和林冠淋溶对穿透雨中养分元素含量有着复杂的影响,所以在测定大气降雨、穿透雨和树干茎流时,很难定量区分它们对林内雨水营养元素的贡献。另一方面,由于林冠截获的干沉降表示系统外部的输入,林冠淋溶主要表示养分在森林系统内部的转移,因而区分它们有非常重要的意义。
川西亚高山森林在调节长江上游地区的水量平衡和保持水土等方面具有重要的作用,但由于地处偏僻的高海拔地区,森林水文观测较为困难,所以有关该区域森林生态系统大气降雨中养分输入情况的研究工作很少。因此,本文以王朗自然保护区紫果云杉(Picea purpurea)林、岷江冷杉(Abies faxoniana)林和白桦(Betula platyphylla)林为研究对象,采用人工观测和自动记录相结合的方法,观测了降雨、穿透雨和茎流特征,分析了大气降雨、穿透雨和茎流的元素含量,并采用Lovett等(1984)和Fan等(2001)提出的复线性回归分析方法区分林冠截获的干沉降和林冠淋溶,量化通过大气降雨的的养分输入量,分析了降雨过程中林冠截获的干沉降和林冠淋溶对林内雨水养分含量的影响,以期为研究川西亚高山森林群落系统的生态学过程提供科学依据。
1 材料与方法 1.1 研究区域自然概况研究地位于四川省平武县王朗国家级自然保护区(103°55′-104°10′E,32°49′-33°02′N),海拔2 300~4 980 m,地处青藏高原-四川盆地的过渡地带,属丹巴-松潘半湿润气候。受季风影响,形成干湿季节差异,干季(当年11月至次年4月)表现为日照强烈、降水少、气候寒冷、空气干燥; 湿季(5—10月)的气候特征为降雨集中、多云雾、日照少、气候暖湿。年平均气温2.5~2.9 ℃(最高气温26.2 ℃,最低气温-17.8 ℃),7月平均气温12.7 ℃,1月平均气温-6.1 ℃,年降雨量805.2 mm,主要植被为针阔混交林和针叶林(巩合德等,2004)。
1.2 群落调查分别在七坪沟、小牧羊场沟、大草坪选择600 m2的标准样地,2个相距最远的样地距离约15 km。利用半球照相仪(尼康鱼眼镜头及尼康数码相机Coolpix 950)照相技术和Hemiview软件分析(Ter Stegee, 1994)、测定研究样地内的叶面积指数。采用每木检尺的方法分别测定样地内树木的基本结构特征,同时调查林下植物的种类和数量。
七坪沟的白桦林由亚高山原始针叶林砍伐后形成,其分布区的自然环境条件与亚高山原始针叶林所处的自然环境条件相一致,只是局部小环境的光照更充足,土壤较干燥,林下灌木种类较少,主要有虎榛子(Ostryopsis davidiana)、悬钩子(Rubus palmatus)等; 草本较多,主要有野青茅(Deyeuxia arundinacea)、白苞蒿(Artemisia lactiflora)等。小牧羊场沟的岷江冷杉林主要树种为岷江冷杉、紫果云杉、并混生少量青杨(Populus cathayana),林下植物灌木较少,主要有忍冬(Lonicera spp.)、茶NFDC5子(Ribes spp.)等; 草本较多,主要有铁线蕨(Adiantum capillus-veneris)、日本羊茅(Festuca japonica)、水金凤(Impatiens noli-tangere)等。大草坪的紫果云杉林主要树种为紫果云杉和圆柏,林下植物灌木较多,主要有缺苞箭竹(Fargesia denudata)、忍冬、悬钩子等,草本较少,主要有羊茅(F. ovina)、茜草(Rubia cordifolia)等。各样地群落学特征见表 1。
|
|

2002年5月在白桦林、岷江冷杉林和紫果云杉林内各安装一套全自动微气候观测仪(Measurement Engineering PTY Ltd., Australia)。观测项目包括空气温湿度(型号:HMP45A)、土壤温度(型号:6507A)、太阳总辐射(型号:SK01-D)、降雨(型号:TB-3A)及风速和风向(型号:034A)。每套微气候观测仪配有3组土壤温度传感器(每组2支传感器)、4支太阳总辐射传感器、1支空气温湿度传感器、2支雨量筒,其中1套配有气压、风速和风向传感器。3组土壤温度传感器分别布置在林冠交接、郁闭林冠和非林冠垂直投影下的土壤中,水平位于土壤5 cm和15 cm层; 4支太阳总辐射传感器布置在不同的林冠下; 1支空气温湿度传感器和2支雨量筒位于离地1.4 m高处(灌木层平均高)。另在林外建有高为11.0 m的观测铁塔,在其顶部安装太阳总辐射、空气温湿度、风速和风向传感器,所有探头用导线与控制器连接,实行24 h连续观测。同时,设置系统扫描间隔为5 s, 数据采集间隔为15 min。采用的微气候数据时间为2002年5月—2003年5月,人工观测时间为2002年7月6日—2003年5月6日,观测期间未收集冰雹、雨夹雪及降雪时的水样。
1.3.1 穿透雨的测定自动测定:每个样地安置2个雨量筒(型号:TB-3A,0.2 mm),可不间断地测定森林内的降雨分配情况。人工测定:把由PVC材料制成的收集面积为0.4 m2(2 m×0.2 m)的集水槽用3%的盐酸连续浸泡24 h,并用蒸馏水至少清洗3次,然后在样地内随机安置5个集水槽,为了避免灌木及草本植物对穿透雨的影响,使集水槽距离地面的高度不低于30 cm,并与地面保持约0.5°的倾角,为了避免枯枝落叶等影响测量结果,降雨前把集水槽内的凋落物等清理干净。集水槽较低的一端底部开口,连接一个体积为7.5 L的塑料桶,最后以5个集水槽的雨量平均数作为穿透雨的量。
1.3.2 大气降雨的测定自动测定:在林内空旷处安置1个雨量筒,其自动测定的值可作为大气降雨的参考数据。人工测定:在林外空旷地约50 m处安置1个集水槽作为对照(CK),测定的值为总降雨量。
1.3.3 茎流的测定根据径级分布,每样地共选择5株标准木观测树干茎流。将直径为1.5 cm大小的聚乙烯塑料管沿中缝剪开一段(剪开的长度取决于树干的大小),然后用大头针将塑料管开口处固定在树干上,再将剪开的塑料管从两边螺旋上升缠绕于树干1圈,固定后,用玻璃胶将接缝处封严,在塑料管的下端接1个2.5 L塑料桶收集树干茎流。每次测定的茎流量的平均值乘以相关径级的树木的数目换算成林分的总茎流量(mL),除以样地面积就是茎流水深(mm)。
1.3.4 林冠截留量的计算利用公式:I=R- T- S计算林冠截留量。其中I为林冠截留量(mm),R为降雨量,T为穿透雨量,S为茎流量。
1.3.5 元素的测定每次降水后,从样地各容器收集到的水分中分别取250 mL的样品,并加入1 mL的HgCl2固定。NO3--N用紫外分光光度法(双光束扫描紫外可见分光光度计,型号UV330,英国LNICAM公司制造)测定,NH4+-N用靛酚蓝比色法(722S分光光度计,上海棱光技术有限公司制造)测定,K、Ca和Mg都用原子吸收分光光度法测定(北京普析通用有限公司PGENERAL-Atomic absorption spectrophotometers); S用硫酸钡比浊法测定; 无机C用双指示剂滴定法测定,有机C用重铬酸钾氧化法测定; 全P用硫酸-高氯酸氧化-钼蓝比色法测定。
1.4 数据分析结合微气候观测仪自动测定的数据,在每个样地中各人工测定了29次大气降雨量、穿透雨量和树干茎流量,并计算得到每个样地每次降雨事件的降雨前干燥时间、降雨强度、降雨持续时间和林冠截留量等,同步分析每次大气降雨、穿透雨和树干茎流的养分元素的浓度(表 2)。每一种元素的林地净输入量可用下式表示:NTF=(TF+SF)-RF。其中NTF表示元素的林地净输入量(kg·hm-2),TF表示元素在穿透雨中的含量,SF表示元素在茎流中的含量,RF表示元素在林外大气降雨中的含量。基于Lovett等(1984)的方法,采用了复线性回归方程区分穿透雨和茎流中的元素净输入量是来源于林冠截获的干沉降(系统外部)还是来源于林冠交换(系统内部)。
|
|

表 2表明:大气降雨中所含各种元素的浓度并不相同,在测定的营养元素中S浓度最高,平均浓度为5.37 mg·L-1,NO3--N的浓度最低,为0.16 mg·L-1(表 2)。不同地区由于年降水量和雨水元素浓度的不同,其各种元素的年输入量也不同。测定地区的各种元素中,S的年输入量最高,达到44.25 kg·hm-2,远高于华北地区6.58 kg·hm-2(李海涛等,1997),这可能是该区的酸沉降导致的(彭培好等,1996)。P随大气降雨输入的年养分量最低为0.4 kg·hm-2,高于湖南会同测定的值0.21 kg·hm-2(田大伦等,2002)。即使在同一地区不同年份和不同地点,各元素的年输入量也不同,K、Ca、Mg的年输入量分别为37.76、38.97、8.05 kg·hm-2,远高于彭培好等(1996)在同一地区(绵阳市新桥镇官司河流域,104°45′—104°48′ E,31°32′—31°37′ N)测定的值(K 2.91、Ca 17.05、Mg 1.66 kg·hm-2),但是N(NH4+-N、NO3--N)和P的年输入量分别为6.44和0.4 kg·hm-2,低于他们的测定结果6.70 kg·hm-2和0.75 kg·hm-2。而大气降雨中C的年输入量为40.57 kg·hm-2。各元素通过大气降雨输入量的大小依次是S>C>Ca>K>Mg>N>P(表 3)。
|
|

表 2表明,大气降雨穿过林冠表面后其化学元素含量发生了很大变化,3种森林群落中穿透雨NO3--N、K、Ca、Mg、C和P浓度都增加,而NH4+-N和S浓度都有所降低,表明二者在经过林冠层被不同程度的吸附,其中白桦林对NH4+-N吸附最明显,原始针叶林对S吸附最为明显。3种森林群落穿透雨中各营养元素浓度明显不同,白桦林穿透雨中的S浓度比紫果云杉林高73.3%,紫果云杉林的Ca和C浓度分别比白桦林高17.1%和61.8%,岷江冷杉林的NO3--N、NH4+-N、Mg和K浓度分别比白桦林高59.6%、126.9%、32.9%和117.7%,岷江冷杉林的P浓度比紫果云杉林高475.0%,这也说明阔叶林和针叶林内雨水营养元素浓度并不相同,而且不同树种对改变降雨养分含量所起的作用也不同,如阔叶林对NH4+-N的吸附大于针叶林,而针叶林林冠对S元素的吸附明显高于阔叶林。营养元素含量的变化是由于雨水淋溶了植物组织上的营养物质、大气降雨淋洗了林冠上的尘埃颗粒以及植物和雨水之间进行的离子交换(Lindberg et al., 1986),另外,林冠截留也会造成穿透雨和茎流营养元素浓度升高(林冠截留占降水量的48.3%)(Laclau et al., 2003)。
茎流的营养元素包括经过林冠和树皮作用后的营养元素(Crockford et al., 1996)。表 2表明:茎流中营养元素浓度明显高于穿透雨中营养元素的浓度,其中K和S元素浓度增加最明显。茎流中营养元素浓度明显升高可能是由两个原因造成的:树皮给大气沉降物提供了良好的接受场所; 茎流量通常较少,只占降雨量的1%~5%,使营养元素出现了浓缩的现象(Fan et al., 2001)。在每个样地中,树干茎流中各营养元素浓度的顺序都和穿透雨一致。表 3表明,尽管树干茎流中养分浓度都较高,但由于茎流量都较小,所以茎流养分输入都很小,在紫果云杉林中甚至可以忽略不计,但由于其养分含量的特殊性,在土壤化学养分输入养分平衡方面有重要意义(Haworth et al., 1995),且茎流直接将营养元素输入到植物根区(Parker, 1983),直接为植物生长提供养分。
2.3 营养元素的林地净输入量特征表 3表明,3种森林群落中NH4+-N、NO3--N和Ca、岷江冷杉林和紫果云杉林内的S及紫果云杉林内的P的林地净输入量都为负值,这与Aboal等(2002)对月桂森林NO3--N和S的研究结果相一致,也与Fan等(2001)对冷杉林的NH4+-N和NO3--N的研究结果一致,这可能由以下原因造成:1)降雨过程中水分的蒸发,而使这些元素留在林冠表面; 2)充分湿润林冠需要的降雨量较高(白桦林1.0 mm,岷江冷杉林1.5 mm,紫果云杉林3.0 mm); 3)研究区域的降雨持续时间长而降雨量小(55.3%的降雨持续时间超过5 h,降雨量<10 mm的降雨占总降雨次数的78.2%),一些移动性差的元素(如Ca)很难冲刷下来; 4)林冠截留率较高(白桦林为19.3%,岷江冷杉林为48.3%,紫果云杉林为34.8%)。这些留在林冠表面的元素可能被随后较强的降雨过程冲刷下来(Neary et al., 1994),也可能被嫩枝和树叶吸收(Kozlowski et al., 1997),或者可能被树干吸收(Mitterhuber et al., 1989); 而其余的元素林地净输入量都很小,低于以前的研究(Moreno et al., 2001; Aboal et al., 2002),说明测定期间林冠截获的干沉降和林冠淋溶对研究的森林群落影响都很小。
因为林冠淋溶主要发生在叶面,并随降雨冲刷下来,所以和雨水相关的因素(如降雨量、降雨持续时间、降雨强度和林冠截留量等)都可能影响林冠淋溶,而降雨间隔时间影响林冠截获的干沉降(Lovett et al., 1984)。为了验证降雨特征对穿透雨和茎流中各种营养元素的影响采用了复线性回归分析,可用下式表示:
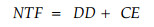
|
式中:NTF为元素的林地净输入量,DD为林冠截获的干沉降,CE为林冠交换(林冠淋溶和降雨中的元素被林冠吸收)。它以如下假设为基础:1)穿透雨中林冠截获的干沉降和雨前干燥时间有关; 2)淋溶量和降雨量有关; 3)回归方程是线性的; 4)林冠截获的干沉降很快而且完全的从林冠洗刷下来(Lovett et al., 1984)。根据Lovett等(1996)对方程的改进和Rea等(2001)对落叶林穿透雨中微量元素的研究,采用了复线性回归模型检验了降雨量、降雨间隔时间、降雨强度、降雨持续时间及大气降雨中元素的浓度等降雨特征(自变量)与林地净输入量(因变量)之间的相关性,发现降雨间隔时间、降雨量和大气降雨中元素的浓度影响穿透雨和茎流中元素的净输入量(α<0.05),回归方程如下:

|
式中:NTFX表示元素X的林地净输入量(mg·m-2),a表示截距,b1、b2、b3表示回归系数,A表示降雨前的干燥时间(降雨间隔时间,h),R表示降雨量(mm),CX表示大气降雨中元素X的浓度(mg·L-1)。
|
|

表 4表明:3种森林群落内降雨量只与S和C元素的林地净输入量明显相关,这与Fan等(2001)对冷杉林内S元素的研究结果一致,在岷江冷杉林内和K也明显相关,说明S与C元素的林地净输入量主要和雨水对植物的淋溶有关,且随降雨量的升高,它们的浓度都显著降低。这可能是由于养分是在降雨过程中阶段被淋溶下来的,而且林冠上的尘埃颗粒也是在降雨过程开始被大量冲洗下来,而对于降雨较大的事件,降雨对养分浓度起了稀释效应。而雨前干燥时间(降雨间隔时间)和NH4+-N、NO3--N、K、Ca、Mg和P等元素林地净输入量高度相关,表明这几种元素可能主要来自于林冠截获的干沉降,很少一部分来自于雨水淋溶。而岷江冷杉林内的K元素林地净输入量与降雨量和降雨间隔时间都明显相关,说明在岷江冷杉林内K与林冠淋溶和林冠截获的干沉降都密切相关。研究发现大气降雨中元素的浓度和3个样地内几乎所有的元素(白桦林内K和Ca元素除外)都显著相关,在国内外的研究中很少见到类似的报道,这表明在降雨过程中这些元素有相当一部分被保留在林冠上或被树叶吸收。验证了降雨持续时间、降雨强度和林冠截留量与各养分元素之间的关系,没有发现它们之间的相关性。
3 结论C、N(NH4+-N和NO3--N)、P、K、Ca、Mg、S等养分随大气降雨输入林地的量分别为40.57、6.44、0.40、38.69、39.92、8.27和44.25 kg·hm-2a-1。
3种森林群落内的S和C可能主要来自于林冠淋溶,而NH4+-N、NO3--N、K(白桦林和紫果云杉林内)、Ca、Mg和P等主要来自于林冠截获的干沉降,岷江冷杉林内K与林冠淋溶和林冠截获的干沉降都密切相关。干沉降和林冠淋溶对所研究的森林群落的影响可能很小。
由于研究期间降雨持续时间长、降雨量小及较高的林冠截留量,可能造成一些元素的林地净输入量为负值。
陈永瑞, 刘允芬, 林耀明, 等. 2003. 江西千烟洲试区木荷阔叶林的水文过程及养分动态研究. 林业科学, 39(4): 64-71. |
樊后保. 2000. 杉木林截留对降水化学的影响. 林业科学, 36(4): 2-8. DOI:10.3321/j.issn:1001-7488.2000.04.001 |
巩合德, 王开运, 杨万勤, 等. 2004. 川西亚高山白桦林穿透雨和茎流特征观测研究. 生态学杂志, 23(4): 1-10. DOI:10.3321/j.issn:1000-4890.2004.04.001 |
黄建辉, 李海涛, 韩兴国, 等. 2000. 暖温带两种针叶林生态系统中茎流和穿透雨的养分特征研究. 植物生态学报, 24(2): 248-251. DOI:10.3321/j.issn:1005-264X.2000.02.021 |
李海涛, 韩兴国, 陈灵芝. 1997. 华北暖温带山地落叶阔叶混交林的茎流研究. 生态学报, 17(4): 371-376. DOI:10.3321/j.issn:1000-0933.1997.04.005 |
彭培好, 王金锡, 胡振宇, 等. 1996. 人工桤柏混交林中降雨对养分物质的淋溶影响. 生态学杂志, 15: 12-15. DOI:10.3321/j.issn:1000-4890.1996.05.001 |
田大伦, 项文化, 杨晚华. 2002. 第2代杉木幼林生态系统水化学特征. 生态学报, 22(6): 859-865. DOI:10.3321/j.issn:1000-0933.2002.06.010 |
Aboal J R, Jiménez M S, Morales D, et al. 2002. Net below canopy fluxes in Canrian laurel forest canopies. Journal of hydrology, 264: 201-212. DOI:10.1016/S0022-1694(02)00095-1 |
Crockford R H, Richardson D P, Sageman R. 1996. Chemistry of rainfall, throughfall and stemflow in a Eucalyptus forest and a pine plantation in south-eastern Australia. 3. Stemflow and total inputs. Hydrological Processes, 10: 25-42. DOI:10.1002/(SICI)1099-1085(199601)10:1<25::AID-HYP297>3.0.CO;2-W |
Fan H B, Wei H. 2001. Estimation of dry deposition and canopy exchange in Chinese fir plantations. Forest Ecology and Management, 147: 99-107. DOI:10.1016/S0378-1127(00)00469-2 |
Haworth K, Mcpherson G R. 1995. Effects of Quercus emoryi trees on precipitation distribution and microclimate in a semiarid savanna. Journal of Arid Environments, 31: 153-170. DOI:10.1006/jare.1995.0057 |
Kozlowski T T, Pallardy S G. 1997. Physiology of woody plants. San Diego: Academic Press.
|
Laclau J P, Ranger J, Bouillet J P, et al. 2003. Nutrient cycling in a clonal stand of Eucalyptus and an adjacent savanna ecosystem in Congo. Forest Ecology and Management, 176: 105-119. DOI:10.1016/S0378-1127(02)00280-3 |
Lindberg S E, Lovett G M, Richter D D, et al. 1986. Atmospheric deposition and canopy interactions of major ions in a forest. Science, 231: 238-244. |
Lindberg S E, Lovett G M. 1985. Field measurements of particle dry deposition rates to foliage and inert surfaces in a forest canopy. Environmental Science Technology, 19: 238-244. DOI:10.1021/es00133a003 |
Lovett G M, Linderberg S E. 1984. Dry deposition and canopy exchange in a mixed oak forest as determined by analysis of throughfall. Journal Applied Ecology, 21: 1013-1028. DOI:10.2307/2405064 |
Lovett G M, Nolan S S, Driscoll C T, et al. 1996. Factors regulating throughfall flux in a New Hampshire forested landscape. Candian Journal of Forest Research, 26: 2134-2144. DOI:10.1139/x26-242 |
Mitterhuber E, Pfanz H, Kaiser W M. 1989. Leaching of solutes by the action of acidic rain: a comparison of efflux from twigs and single needles of Picea abies (L. Karst). Plant and Cell Environment, 12: 93-100. DOI:10.1111/j.1365-3040.1989.tb01920.x |
Moreno G, Gallardo J F, Bussotti F. 2001. Canopy modification of atmospheric deposition in oligotrophic Quercus pyrenaica forests of an unpolluted region (central-western Spain). Forest Ecology and Manegement, 149: 47-60. DOI:10.1016/S0378-1127(00)00544-2 |
Neary A J, Gizyn W I. 1994. Throughfall and stemflow chemistry under deciduous and coniferous forest canopies in south-central Ontario. Canadian Journal of Forest Research, 24: 1089-1100. DOI:10.1139/x94-145 |
Parker G G. 1983. Throughfall and stemflow in the forest nutrient cycle. Advances in Ecological Research, 13: 57-134. DOI:10.1016/S0065-2504(08)60108-7 |
Rea A W, Lindberg S E, Keeler G J. 2001. Dry deposition and foliar leaching of mercury and selected trace elements in deciduous forest throughfall. Atmospheric Environment, 35: 3453-3462. DOI:10.1016/S1352-2310(01)00133-9 |
Rodrigo A, Àvila A, Rodà F. 2003. The chemistry of precipitation, throughfall and stemflow in two holm oak (Quercus ilex L.) forests under a contrasted pollution environment in NE Spain. The Science of The Total Environment, 305: 195-205. |
Ter Stegee H. 1994.HEMIPHOT: a programme to analyse vegetation indices, light and light quality from hemispherical photographs. Tropenbos Documents 3. The Tropenbos Foundation, Wageningen, The Netherlands
|