 2005, Vol. 41
2005, Vol. 41文章信息
- 周志春, 谢钰容, 金国庆, 陈跃, 宋振英.
- Zhou Zhichun, Xie Yurong, Jin Guoqing, Chen Yue, Song Zhenying.
- 马尾松种源磷效率研究
- Study on Phosphorus Efficiency of Different Provenances of Pinus massoniana
- 林业科学, 2005, 41(4): 25-30.
- Scientia Silvae Sinicae, 2005, 41(4): 25-30.
-
文章历史
- 收稿日期:2003-09-15
-
作者相关文章


2. 福建省南纸股份有限公司 南平 353000
2. Fujian Nanping Paper Co. Ltd. Nanping 353000
地壳中全磷含量高达0.12%(Meng et al., 1982),但可供植物吸收利用的有效磷含量却很低,如华南地区低磷酸性红壤的有效磷含量为2 mg·kg-1 (李庆逵, 1985)。传统上常通过改良土壤或施肥措施解决有效磷的缺乏。然而大量施用磷肥不仅提高生产成本,而且造成土壤磷素异常累积、水体富营养化等生态问题(曹靖等, 2000)。研究发现,不同植物对土壤磷的吸收和利用存在显著差异,例如杂草的富磷能力通常较高(Gupta et al., 1984)。这些差异主要体现在磷素吸收和积累量(Clark et al., 1974; Baker et al., 1970)、磷素代谢特性(Fageria et al., 1988; Yan et al., 1995)以及干物质生产力等方面(高守疆等, 1989)。近年来,植物磷效率遗传改良研究受到人们的关注。培育有效活化和吸收利用缺磷土壤中潜在磷素(如闭蓄态磷)的植物新品种,可从根本上解决农林业上土壤有效磷“遗传学缺乏”的难题。
马尾松(Pinus massoniana)是我国南方主要的乡土造林和工业原料树种,分布广泛,适应性强,生长迅速。自20世纪60年代开展马尾松种源试验以来,其遗传改良研究取得了重大进展,为不同地理区域选育出一批速生优质的林木新品种(周志春等, 1997)。然而马尾松已有育种方案未考虑我国森林土壤地力不断衰退,缺磷少氮严重的现状,在缺磷的林地上应用这些育成的品种将难达到预期效果。为此,本项目组启动了马尾松耐低磷育种的研究。通过设计不同磷水平处理的水培和土培试验,已从磷素吸收动力学参数、根际有机酸分泌物和酸性磷酸酶活性、根系形态建成和干物质分配格局等方面初步阐明了马尾松不同种源对低磷胁迫的反应差异和适应机制(谢钰容等, 2003; 2004a; 2004b; 2005)。本文继续利用不同磷水平处理的盆栽试验,开展马尾松种源磷效率的研究,同时确立磷效率的特异性指标,为培育和筛选有效利用土壤磷素的马尾松优良基因型提供理论依据。
1 材料与方法 1.1 材料来源选择浙江淳安、福建武平、江西崇义、广东信宜和广西岑溪5个马尾松种源参试,这些种源分别地处我国亚热带的南、中和北带地区,代表了不同的地理气候条件,对低磷胁迫和磷肥的遗传反应存在显著差异(周志春等, 2003)。盆栽试验基质取自中国林科院亚林所虎山(浙江省富阳市)的贫瘠缺磷酸性红壤,其全磷和有效磷含量分别为0.94 g·kg-1和1.87 mg·kg-1,pH4.55。供试土壤的其他性质参见文献(谢钰容等,2003)。
1.2 盆栽试验基质土壤经风干过筛后,将过磷酸钙、珍珠岩一起与土壤混合均匀后装盆。2002年3月在中国林科院亚林所开展不同种源的磷肥盆栽试验。设置低磷(不施P,P0)、中磷(土壤中过磷酸钙含量为1.0 g·kg-1,P1)和高磷(土壤中过磷酸钙含量为2.0 g· kg-1,P2)3种磷水平处理,3次重复,24盆小区。营养杯直径8 cm、高10 cm。试验用磷肥为浙江绍兴产,含P2O5 12.4%。N、K等其他养分保持正常水平。盆栽试验为露天栽培,每营养杯移栽芽苗2株,至苗木生长正常后保留1株,正常管理直到收获。
1.3 试验收获和测定盆栽试验苗于2002年10月收获。不同磷水平下每种源选取5株平均大小的苗木,先量测苗高生长和根系形态参数(谢钰容等,2004b),然后将幼苗分成根、茎、叶3部分,经105 ℃杀青30 min,79 ℃烘干至恒重。用浓H2SO4-H2O2消煮—钼锑抗比色法测定根、茎、叶各部分的含磷量(中国土壤学会农业化学专业委员会, 1983)。
1.4 磷效率的计算和数据分析磷效率、吸收效率和利用效率分别按照如下公式计算(曹靖等, 2000):
磷效率=低磷胁迫下干物质积累量/(中)高磷条件下干物质积累量×100%;磷素吸收效率=磷吸收量/株;磷素利用效率=干物质积累量/磷吸收量。
用SAS统计软件的ANOVA程序按种源、磷肥水平分别进行2因素和单因素的方差分析,以检验种源、磷肥效应。采用简单相关分析估算低磷水平下马尾松种源磷效率与其子性状的相关系数。
2 结果与分析 2.1 马尾松种源干物质积累及磷效率的差异方差分析发现,盆栽条件下马尾松干物质的积累存在显著的种源效应和磷肥效应。磷素营养的改善将使马尾松种源的干物质积累量显著提高,其增幅为40.5%~133.7%(中磷)与39.4% ~193.7%(高磷)(表 1)。在低磷胁迫下,福建武平种源的干物质生产能力最强,较其他参试种源高出69.2%~95.7%,表明该种源能较好地适应低磷环境。比较不同磷处理水平下种源的干物质积累量,还可以发现,福建武平种源对磷肥的敏感性较低,广东信宜和浙江淳安种源次之,而广西岑溪和江西崇义种源对磷肥投入可获得较高的增益。
|
|
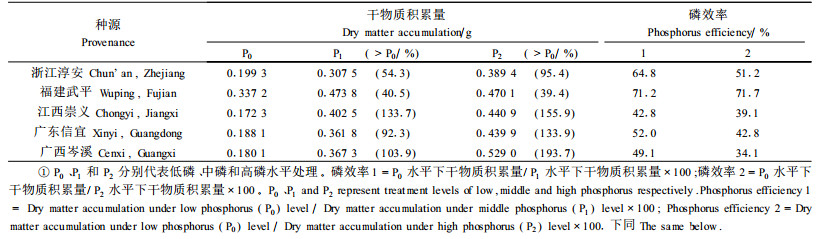
磷效率表示为磷胁迫条件下的干物质积累量与正常磷水平时的差异,体现了植物对低磷逆境适应能力的强弱,能较好地反映植物在磷胁迫下的生长情况。比较发现,地处武夷山脉南端的福建武平种源具有最高的磷效率(中磷71.2%、高磷71.7%),分布于较北缘的浙江淳安种源次之。在参试的3个南方种源中,广东信宜种源的磷效率也相对较高,分别达52.0%(中磷)和42.8%(高磷),广西岑溪和江西崇义种源的磷效率则较低。马尾松种源磷效率特性的不同,意味其对磷肥反应式样具有差异性。福建武平和广东信宜等种源在低磷胁迫下能获得相对于较高磷素水平(中磷和高磷)较高比例的干物质积累量,这一结果与大田试验和水培试验相吻合(周志春等, 2003;谢钰容等, 2003)。
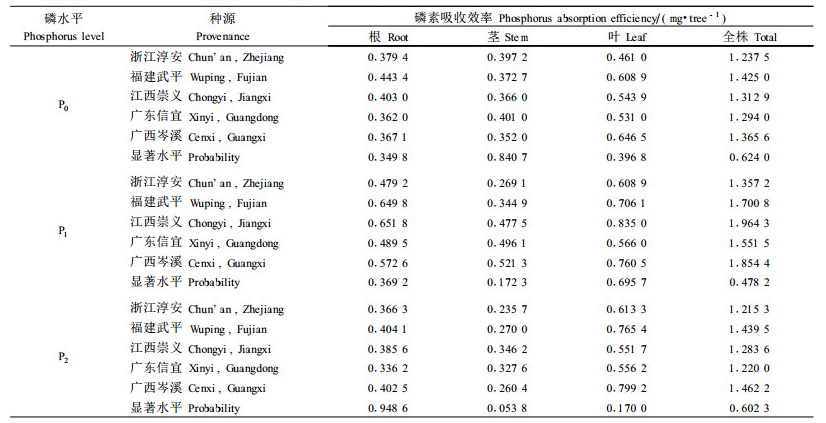
2.2 不同磷水平下马尾松种源的磷素吸收效率统计分析表明,马尾松对磷素的吸收与磷素的环境有关,不管是以根、茎和叶为单元,还是以整个植株为计算单元,其磷素吸收效率在磷处理水平间都达到极显著的差异(p=0.096 0~0.000 1)。与低磷处理比较,中磷水平下多数种源根、茎、叶和全株的磷素吸收绝对量显著增加,但在高磷水平下,除针叶外,种源磷素的吸收量变化较小(表 2)。在相应的磷素水平下,不同种源的磷素吸收量虽存在一定的差异,却未达到统计学上的显著水平,因此单纯从磷素吸收效率上难以区分耐低磷型种源和磷高效型种源。研究还发现,低磷水平下福建武平种源根系和全株吸收的磷量较多,分别为0.443 4 mg·株-1和1.425 0 mg·株-1。
|
|
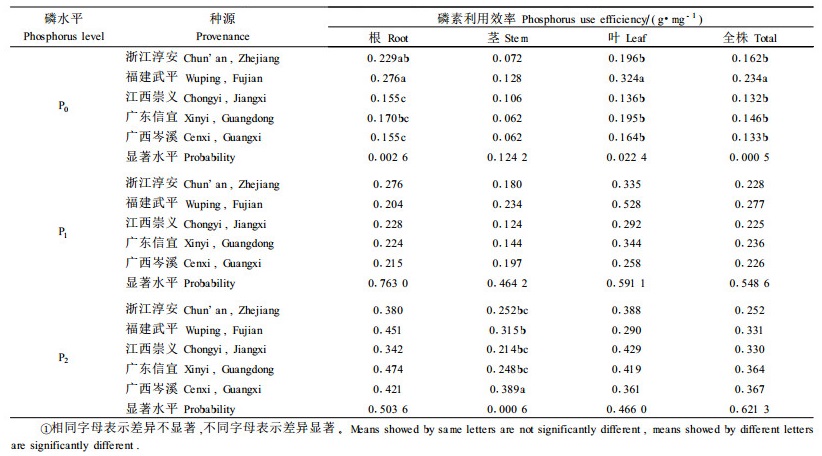
磷素利用效率是指植物吸收单位磷量所产生的干物质量,干物质量越高,表明其对吸收磷素的利用越充分。方差分析表明,马尾松根、茎、叶和全株的磷素利用效率存在显著的磷肥效应(p < 0.000 1)和种源效应(p=0.051~0.098)。表 3按磷处理水平列出了各种源根、茎、叶和全株磷素利用效率的估算值。研究发现,低磷胁迫下,除茎外,根、叶和全株的磷素利用效率在种源间的差异都达到了极显著水平。多重对比分析发现福建武平种源的磷素利用效率最高,其根、叶和全株估算值分别为0.276、0.324和0.234 g·mg -1。这一结果意味着福建武平种源能较好地适应低磷胁迫并具有较高的磷效率,一方面因其磷素吸收能力略强,另一方面因其对已获得磷素的利用效率较高。表 3还表明,随着土壤磷素营养的改善,种源根、茎、叶和全株的磷素利用效率都显著提高,但种源间的差异却在逐渐减小。中高磷水平下,虽然不同种源磷素利用效率的绝对值存在较大的差异,但在统计学上未达到显著水平。这里例外的是,高磷水平下茎的磷素利用效率具有显著的种源效应,其原因不明。
|
|
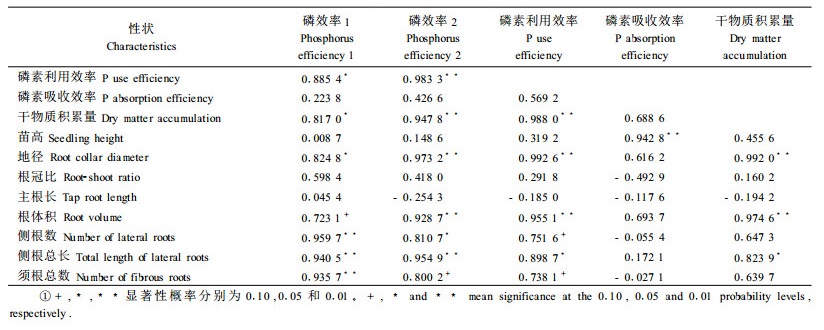
磷效率反映的是植物在介质有效磷浓度较低时正常生长的能力。这里进一步研究种源磷效率(包括磷素利用效率和吸收效率)与苗木生长、干物质积累、根系参数的相关性,以确立马尾松种源磷效率的特异性指标。表 4结果显示,低磷胁迫下种源磷效率与磷素利用效率呈显著的正相关,而与磷素吸收效率相关性较小,这意味着种源有效利用较低磷素浓度的能力可较好地反映其磷效率的高低。盆栽试验条件下,种源磷效率与苗高的相关性较小,而与地径有关。种源磷效率与根系参数的相关结果揭示,低磷胁迫下一些根系形态特征可很好地预测马尾松种源磷效率和磷素利用效率的大小,磷效率和磷素利用效率高的种源侧根发达、须根数量多,进而具有较高的干物质生产能力。根体积、侧根数、侧根总长和须根总数等根系参数可作为低磷胁迫下马尾松磷效率的特异性指标。由于盆栽条件下苗木主根的生长受到育苗容器大小的制约,这里未发现种源主根长、根冠比与磷效率间的显著相关性。
|
|
植物中普遍存在着磷效率特性基因型之间的差异。在同样的低有效磷土壤中,一些种类的植物可以正常生长,而另一些则生长严重受阻,甚至死亡。不同生态区域都存在着一些突出的高磷效率植物种类,并形成各种应对低磷胁迫的机制。不同植物基因型的磷效率差异非常大,如在其他养分供应正常但土壤有效磷较低的条件下,来自不同基因库的菜豆(Phaseolus vulgaris)基因型苗期生物量和籽粒产量差异分别达4.2倍和2.4倍(严小龙等, 1997)。对植物基因型磷效率特性的评价有不同的指标,归纳起来主要有基因型的磷素吸收量、植株干物质的积累和磷素代谢特性等,这些指标在不同的研究中被单独或被联合采用(高守疆等, 1989),在不同植物上有所侧重。如低磷水平下冠部磷素吸收量可以简便有效地衡量大豆(Glycine max)基因型的磷效率差异(童学军等, 2000)。寻求磷效率的特异性指标,尤其是一些简单易测的形态和生理指标特征,在耐低磷植物育种中具有重要意义。
本研究布设了3种磷水平处理的盆栽试验,以揭示马尾松种源磷效率及磷素吸收和利用效率特征的差异,确立马尾松种源磷效率的特异性指标。结果表明,马尾松不同种源的磷效率差异巨大。来自武夷山脉南端福建武平种源的磷效率最高,而来自云开大山和南岭山脉的广西岑溪、江西崇义等南部种源的磷效率则相对较低。福建武平种源所处的闽西地区是著名的红土地,其土壤的有效磷含量非常低。长期适应缺磷土壤的结果使得这里的种源具有耐低磷特性。来自这一地区的种源适应性好,稳定性高(周志春等, 1998),对土壤磷的活化和富集作用强烈(周志春等, 2003)。云开大山和南岭山脉是马尾松的主要中心产区,其自然环境条件优越,来自该区域的种源速生丰产性显著,但稳定性低、遗传可塑性大,对磷肥投入敏感。然而,广东信宜种源不同于该区域的其他种源,磷效率相对较高,可认为这也是一个能较好适应低磷胁迫的优良种源。
低磷胁迫下马尾松家系间的磷素吸收效率和利用效率也差异显著,但发现家系的磷效率与其在低磷水平下的磷素吸收效率相关性较小,而与磷素利用效率呈显著的正相关,磷素利用效率而非磷素吸收效率是造成马尾松磷效率和干物质家系差异的主要原因,这类似李继云等(2000)对小麦(Tritium aestivum)品种磷效率的研究结果。
植物磷效率的高低取决于其对土壤磷的活化、有效吸收和利用的能力或效率。作者已从根际土壤营养(周志春等, 2003)、根系有机酸分泌物特性和分泌性酸性磷酸酶活性差异(谢钰容等, 2004a; 2005)等研究比较了马尾松不同种源活化土壤难溶态磷的能力。这里则主要从磷素吸收效率和磷素利用效率两方面阐明马尾松种源磷效率的特征差异。研究发现,低磷胁迫下马尾松参试种源的磷素利用效率差异很大,福建武平种源的测定值最高,而江西崇义种源的测定值最低,两者相差近1倍。相关分析表明,磷素利用效率高的种源,其磷效率和干物质积累量也较高,这说明低磷条件下的磷素利用效率可用以反映种源的磷效率大小。虽然观测到福建武平种源在低磷水平下的磷素吸收效率相对较高,但与其他种源未达到显著差异,仅依据磷素吸收效率还难以把握种源的磷效率特征。这一结果说明,磷素利用效率而非磷素吸收效率是造成马尾松磷效率和干物质种源差异的主要原因。李继云等(2000)在中低磷水平下研究小麦品种磷吸收和利用效率与生物产量和籽粒产量关系时也获得类似的结论。这里的研究结果有异于水培实验(谢钰容等, 2003)。在水培试验中,发现低磷条件下马尾松种源的磷素吸收动力学参数差异较大,其中福建武平和广东信宜2个种源具有较大的Imax(最大吸收速率)和较小的Km(1/2Imax时介质的离子浓度)和Cmin(吸收速率为0时的介质离子浓度)。其实,马尾松在水培和土培2种试验体系中对低浓度磷的吸收方式是不同的,水培试验结果不能代表土培的真实情况。
衡量根系大小和形态的一些指标是与磷效率密切相关的子性状,受较强的遗传控制,可以作为磷效率育种的重要评价依据(严小龙等, 1997)。低磷条件下马尾松种源磷效率与主要根系参数的相关结果表明,磷效率与根体积、侧根数、侧根总长和须根总数等根系参数呈显著的正相关,而与根冠比和主根长的相关性较小。结合已有的水培和土培试验结果,初步认为根体积、侧根数、侧根总长和须根总数等根系参数及根系有机酸分泌物含量、酸性磷酸酶活性等生理指标可作为低磷胁迫下马尾松种源磷效率的特异性指标。
曹靖, 张福锁. 2000. 低磷条件下不同基因型小麦幼苗对磷的吸收和利用效率及水分的影响. 植物生态学报, 24(6): 731-735. DOI:10.3321/j.issn:1005-264X.2000.06.015 |
高守疆, 陈升枢, 李明启. 1989. 不同磷营养水平对烟草叶片光合作用和光呼吸的影响. 植物生理学报, 13(3): 281-287. |
李继云, 孙建华, 刘全友, 等. 2000. 不同小麦品种的根系生理特性、磷的吸收利用效率对产量影响的研究. 西北植物学报, 20(4): 503-510. DOI:10.3321/j.issn:1000-4025.2000.04.002 |
李庆逵主编. 1985.中国红壤.北京:科学出版社, 145-146
|
童学军, 卢永根, 严小龙. 2000. 广东大豆地方种质磷效率特性研究. 中国油料作物学报, 22(4): 49-53. |
谢钰容, 周志春, 金国庆, 等. 2003. 马尾松不同种源磷素吸收动力学特征. 林业科学研究, 16(5): 548-553. DOI:10.3321/j.issn:1001-1498.2003.05.005 |
谢钰容, 周志春, 金国庆, 等. 2004a. 低磷胁迫下马尾松种源根系分泌的主要有机酸研究. 林业科学研究, 17(亚林所所庆专刊): 45-49. |
谢钰容, 周志春, 金国庆, 等. 2004b. 低P胁迫对马尾松不同种源根系形态和干物质分配的影响. 林业科学研究, 17(3): 272-278. |
谢钰容, 周志春, 廖国华, 等. 2005. 磷饥饿诱导下马尾松种源酸性磷酸酶活性差异. 林业科学, 41(3): 58-62. DOI:10.3321/j.issn:1001-7488.2005.03.010 |
严小龙, 张福锁. 1997. 植物营养遗传学. 北京: 中国农业出版社, 44-50.
|
中国土壤学会农业化学专业委员会编. 1983.土壤农业化学常规分析方法.北京: 科学出版社
|
周志春, 秦国峰, 李光荣, 等. 1997. 马尾松遗传改良的成就、问题和思考. 林业科学研究, 10(4): 435-442. DOI:10.3321/j.issn:1001-1498.1997.04.017 |
周志春, 金国庆. 1998. 马尾松不同产地的遗传稳定性和生态学基础. 南京林业大学学报, 22(3): 75-80. |
周志春, 谢钰容, 金国庆, 等. 2003. 马尾松种源对施肥的遗传反应及根际土壤营养差异. 林业科学, 39(6): 62-67. DOI:10.3321/j.issn:1001-7488.2003.06.010 |
Baker D E, Jarrell A E, Marshall L E, et al. 1970. Phosphorus uptake from soil by corn hybrids selected for high and low phosphorus accumulation. Agronomy Journal, 62: 103-106. DOI:10.2134/agronj1970.00021962006200010033x |
Clark R B, Brown J C. 1974. Differential phosphorous uptake by phosphorus-stressed corn inbred. Crop Science, 14: 505-508. DOI:10.2135/cropsci1974.0011183X001400040004x |
Fageria N K, Wright R J, Baligar V C. 1988. Rice cultivar evaluation for phosphorus efficiency. Plant and Soil, 111: 105-109. DOI:10.1007/BF02182043 |
Gupta U C, Macleod J A. 1984. Effect of various sources of phosphorus on yield and P concentration of cereals and forages. Canadian Journal of Soil Science, 64: 403-410. DOI:10.4141/cjss84-041 |
Meng K, Kirkby E A. 1982.Principles of Plant Nutrition. 3rd Ed. International Potash Institute, Bern, Switzerland, 389-409
|
Yan X, Lynch J P, Beebe S E. 1995. Phosphorus efficiency in common bean genotypes in contrasting soil types Ⅰ.Vegetative Response. Corp Science, 35: 1086-1093. |