 2005, Vol. 41
2005, Vol. 41文章信息
- 王莹莹, 左金淼, 刘家冈.
- Wang Yingying, Zuo Jinmiao, Liu Jiagang.
- 以态势理论为基础的更新生态位测度研究
- Study on Regeneration Niche Metrics Based on Ecostate-Ecorole Theory
- 林业科学, 2005, 41(4): 20-24.
- Scientia Silvae Sinicae, 2005, 41(4): 20-24.
-
文章历史
- 收稿日期:2003-10-27
-
作者相关文章


2. 北京林业大学基础科学与技术学院 北京 100083
2. College of Basic Science and Technique, Beijing Forestry University Beijing 100083
生态位理论(niche theory)在种间关系、群落结构、种的多样性及种群进化研究中已被广泛应用,是现代生态学中一个非常重要的概念。自Grinnell(1917)最初明确提出生态位的概念以来,就如何确切地定义生态位及如何计算生态位的测度,一直是生态学界尚未定论的问题。其中最具代表性的当推Grinnell, Elton(1927)和Hutchinson (1957)提出的“空间生态位”、“功能生态位”和“多维超体积生态位”。我国学者也从不同的角度分别给生态位下了定义(王刚等,1984;刘建国,1987;刘建国等,1990;张光明等,1997)。对于生态位测度的研究主要集中在生态位宽度和生态位重叠这2个指标的估算与分析上。在森林群落研究中,生态位宽度计算较多使用的是Levins公式中的Shannon-Wiener指数:
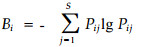
|
(1) |
式中:Bi为i物种的生态位宽度; S为资源单位数,以每个样方作为一资源位; Pij 为i物种在第j资源单位中的重要值百分率。
比较客观反映种群生态位重叠的是Pianka(1975)公式:
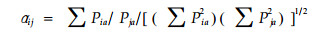
|
(2) |
式中:aij=aji,为种i和种j的生态位重叠值; Pia和Pja分别代表种i和种j中利用资源状态a的重要值百分率。
应用较多、能客观反映种群生态位的还有王刚等(1984;1995)的公式,他将生态位定义为从环境状态集合A={x/x=(x1, …, xn)}到物种Y的密度集合的一个映射f (x1, …, xn)。在此定义下提出了生态位测度的改进公式,不仅具有几何上的直观性,便于理解,而且在实际应用中也能很客观地反映出种群间的生态位重叠以及重叠关系的变化。
更新生态位(regeneration niche)最早由Grubb(1977)提出,主要指植物在有性繁殖和无性增殖过程中对生境条件、空间等生态因子的要求及适应,是植物更新和生态位研究的一个重要方面。有关以林隙作为物种更新生态位的研究,国外有些学者进行了尝试,伐倒树木,在森林中制造不同大小级的林隙,对其植物更新进行研究。我国有关更新生态位研究的报道还很少,如臧润国等(2001)对海南岛霸王岭热带山地雨林林隙更新生态位进行了研究,分析了树种在不同大小、不同年龄级林隙内生态位宽度的变化。
更新生态位本身是一个动态的概念,但一直以来对更新生态位的测量都延用生态位的测度公式,没有考虑到更新生态位的动态性,忽略了生态位的潜在形式和非存在形式。因此,本文提出的更新生态位测度改进公式综合反映了各树种在林隙更新中的相对地位,不仅完善了其生态学意义,而且发展了生态位测度的时间因子的动态特性,为更新生态位的研究提供了新的方法。即根据朱春全(1997)提出的生态位态势理论,改进了更新生态位测度公式。认为在林隙内,大乔木代表该物种在林隙中的当前地位,可以称为物种的当前态; 小径木经过种间竞争和种内竞争,有一部分会长成大乔木,反映了林隙更新的趋势,体现了其对森林生态系统的影响力和支配力,可以称为物种的未来态。因此,分别以大乔木和小径木的生态位作为林隙更新生态位的态和势,在对更新生态位测度公式进行改进的基础上,对长白山阔叶红松林主要乔木树种的更新生态位进行了研究。
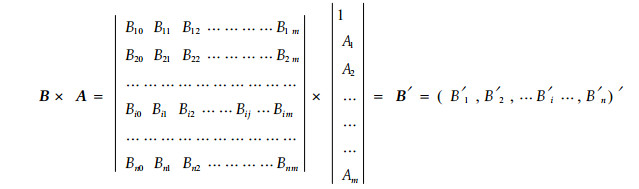
1 更新生态位测度公式改进由态势理论出发可以构建一个更新生态位宽度矩阵模型。计算不同物种的更新生态位,首先构建一个各物种的态势生态位矩阵B, 即把各物种的小径木分成m个径级,再用Levins公式分别计算大乔木和各径级小径木的生态位宽度Bij,由此构成一个n个物种的综合态势矩阵。
|
式中:i=1,2,…,n,j=0, 1, …, m, 其中,B10,B20,…Bn0为n个物种大乔木的生态位宽度; Bi1,Bi2,…Bim为i物种m个径级的生态位宽度。矩阵A为物种各径级态势对该物种更新生态位宽度的影响系数,Aj为物种第j个径级的小径木长成大乔木的成活率,并假设相同径级的各物种长成大乔木的成活率都相同; 得到B′即为n个物种的更新生态位宽度矩阵。
本文由于试验条件限制,只取一个径级的小径木对更新生态位进行了简单计算。即取3 cm < DBH < 10 cm的小径木作为更新生态位的势,取DBH>10 cm的乔木为更新生态位的态,则B′ i=BiB+Ai×BiS,为物种i的绝对更新生态位。为方便比较,取其与该生态系统中其他物种绝对生态位之和的比值即为该物种的相对生态位,如下公式所示:
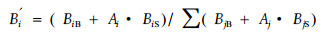
|
(3) |
式中:i,j=1,2,…,n,B′i为改进后的i物种的更新生态位宽度; BiB、BjB为物种i、j大乔木的Levins生态位宽度值; BiS、BjS为物种i、j小径木的Levins生态位宽度值; Ai、Aj为物种i、j在阔叶红松林群落中的重要值百分比,相当于小径木成活率的百分比,为小径木对种群生态位宽度的影响系数。
更新生态位重叠公式改进:
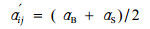
|
(4) |
式中:a′ij为改进后的更新生态位重叠值; aB和aS由Pianka公式(2)计算得到,分别为大乔木和小径木的种间生态位重叠值。此式不仅反映了两物种i、j在当前状态下生态位的相似性,还考虑了被下一代新个体替代后,可能的生态位重叠值,即综合考虑了两物种i、j更新过程的态势特性。
2 研究地区的自然条件和研究方法 2.1 研究地区概况研究地在长白山自然保护区管理局后山1号样地(128°6′E,42°25′N)的阔叶红松混交林,海拔740 m,平缓地,土壤为发育在火山灰上的暗棕壤,土层厚度>100 cm。年均气温为0.9~3.9 ℃,年均降雨量632.8~782.4 mm,森林群落可明显分为4~5层。主要树种有红松(Pinus koraiensis)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、五角槭(Acer mono)、青楷槭(A. tegmentosum)、柠筋槭(A. triflorum)、白牛槭(A. mandshurica)、春榆(Ulmus japonica)、假色槭(A. psudo-sieboldianum)、花楷槭(A. ukurunduense)、千金榆(Carpinus cordata)、蒙古栎(Quercus mongolica)、山槐(Maackia amurensis)、黄檗(Phellodendron amurense)等, 主要下木树种有毛榛子(Corylus mandshurica)、暴马丁香(Syringa reticulata var. mandshurica)、刺五加(Acanthopanax senticosus)等。
2.2 资料收集与整理采用样方法获取长白山阔叶红松林林隙更新生态位测度的原始数据,在1号样地内选择了7个20 m×20 m的林隙样方,调查每个样方的郁闭度、样方内扩展林隙面积、冠林隙面积及主要乔木树种(DBH≥3 cm)的胸径、枝下高、树高,并记录每棵树的相对位置。根据各树种在不同样方中的个体数、胸径及其出现频率,分别算出其在各样方内的相对密度(DR)、相对基盖度(PR)和相对频度(FR),进而得出各树种的重要值:
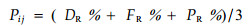
|
(5) |
阔叶红松林树种重要值数据来源于孙中伟等(1995)椴树阔叶红松林群落的数量特征(见表 1)。
|
|
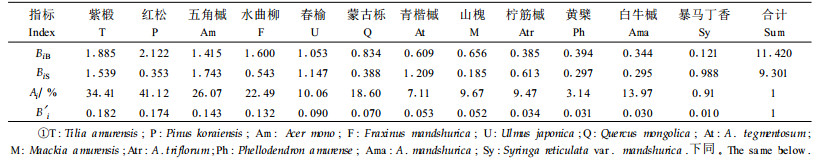
生态位宽度是指现实生态位超体积的限度,反映种群对资源获取和利用状况的尺度。生态位宽度越大,表明该物种的特化程度越小,对环境的适应能力也越强,更倾向于泛化物种。对长白山阔叶红松林主要乔木树种更新生态位宽度的Shannon-Wiener指数和改进公式进行比较分析,并分析了林隙早期演替和晚期演替的更新生态位。
从表 1可见,Shannon-Wiener指数计算得大乔木生态位宽度:红松>紫椴>水曲柳>五角槭>春榆>蒙古栎>山槐>青楷槭>黄檗>柠筋槭>白牛槭>暴马丁香。其中红松更新生态位宽度最大,为2. 122;其次为紫椴、水曲柳、五角槭、春榆,更新生态位宽度值较接近,都大于1;暴马丁香生态位宽度最小,只有0.121。基本与阔叶红松林物种重要值百分比大小顺序相符,反映了林隙植被演替的成熟态。小径木生态位宽度:五角槭>紫椴>青楷槭>春榆>暴马丁香>柠筋槭>水曲柳>蒙古栎>红松>黄檗>白牛槭>山槐。其生态位宽度与大乔木生态位宽度相差很大,反映了林隙更新的趋势。其中五角槭生态位宽度最大,为1.743;其次紫椴、青楷槭、暴马丁香等一些阔叶树在更新过程中优势较大; 而红松则更新较慢,生态位宽度只有0.353,与野外观察结果一致,符合阔叶红松林生态学规律。而改进公式计算得紫椴的相对更新生态位宽度最大,其次为红松、五角槭、水曲柳和春榆,暴马丁香更新生态位最小。与大乔木的生态位宽度相比,能更好地反映阔叶树在林隙更新中的优势。
|
|
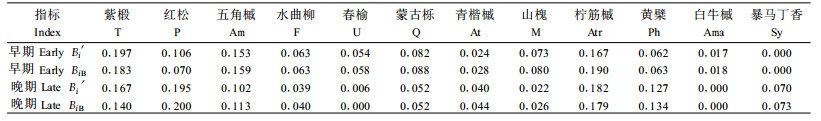
人为区分样方中郁闭度较小的4个林隙样方和郁闭度较大的3个林隙样方为林隙更新演替的早期和晚期,分别计算其更新生态位宽度,并进行比较分析。由表 2可见,对于阔叶红松林形成林冠空隙的初期,更新生态位宽度顺序为:紫椴>水曲柳>红松>五角槭>蒙古栎>山槐>青楷槭>春榆>柠筋槭>黄檗>暴马丁香>白牛槭。按Shannon-Wiener指数计算大乔木生态位宽度得:红松最大,其次为紫椴、水曲柳、蒙古栎、山槐等,白牛槭最小。在林隙形成的早期,紫椴、水曲柳等阳性先锋树种首先侵入,占据较宽的生态位。可见改进后的更新生态位公式能准确地反映这一点。林隙演替晚期改进公式计算得更新生态位宽度顺序为:五角槭>红松>紫椴>春榆>水曲柳>白牛槭>蒙古栎>黄檗>青楷槭>山槐>柠筋槭>暴马丁香。随着林隙年龄的增加,郁闭度增大,树种间的竞争逐渐增大,阳性树种的生长受到限制,五角槭、白牛槭等阴性树种逐渐增多。分别用Shannon-Wiener指数与改进公式计算,得到的大乔木生态位宽度基本一致,因为此时林隙更新基本完成,趋向成熟阶段。林隙演替早期红松相对更新生态位宽度为0.167,演替晚期为0.182,其更新优势比早期大。演替早期紫椴、水曲柳、山槐和柠筋槭的环境适应性更强,而演替晚期随着环境的改变,五角槭、春榆、黄檗、白牛槭的更新优势相对较大。
3.2 阔叶红松林中主要乔木树种的更新生态位重叠生态位重叠是表明不同物种利用生态资源能力异同性的一个指标,生态位重叠值越大,表明2个物种利用资源的能力越相似。本文采用Pianka公式分别计算大乔木和小径木的物种间的生态位重叠值,能客观地反映出种群之间对资源利用或生态适应的相似性。再利用公式(4)计算林隙的更新生态位的种间重叠值。如表 3所示,红松和青楷槭,春榆和水曲柳、假色槭、五角槭,紫椴和柠筋槭,水曲柳和蒙古栎、五角槭,柠筋槭和青楷槭,青楷槭和假色槭,蒙古栎和假色槭,五角槭和假色槭间的生态位重叠都大于0.5,说明它们之间的种间竞争较大。但是红松和阔叶树之间竞争相对较弱,只与青楷槭间的生态位重叠大于0.5,其他都是阔叶树间的竞争,反映出在阔叶红松林群落结构中有利于红松生长更新。
|
|
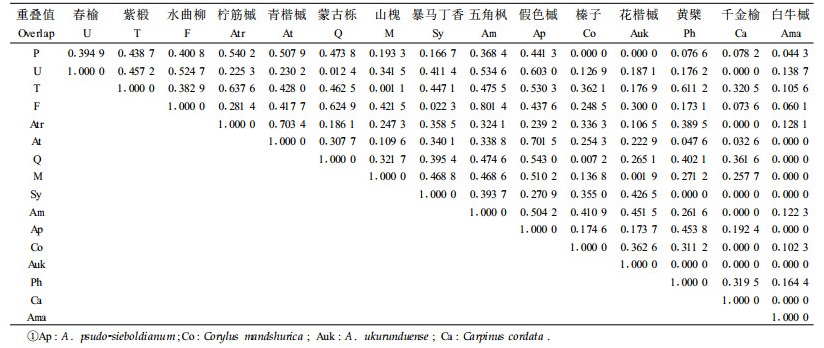
对于长白山阔叶红松林林隙,紫椴的更新生态位宽度最大,其次为红松、五角槭、水曲柳、春榆、蒙古栎等,白牛槭和暴马丁香的更新生态位宽度最小,基本与林下种群的重要值百分比相一致,说明该阔叶红松林种群结构相对稳定。更新生态位宽度较大的物种在林隙中的生态适应性较强、种群优势度较大,将来在林下也将占据较高的比例; 反之,白牛槭和暴马丁香等更新生态位较小的物种在林隙中的生态适应性较弱,将来在整个森林中也将占据较小的比例。
林冠空隙形成后, 空隙内环境条件发生了很大程度的变化, 不同树种对此有不同的反应, 阳性先锋树种(pioneer species)不断侵入, 出现频度较高,占据较宽的生态位,阴性树种自然死亡。林隙演替早期紫椴、水曲柳、山槐和柠筋槭的环境适应性更强,改进公式能很好地反映这一点。随着林冠空隙形成年龄的增加, 不同树种的不同个体在对林冠空隙资源的利用和竞争中形成了各自生态位的分化, 树种的特征也会在生存竞争中随林冠空隙年龄的增加而改变, 直至林冠空隙被填充完毕后, 进入林冠层的只有少数树种的部分个体。演替晚期物种间的竞争逐渐增大,阳性树种的生长逐渐受到限制,阴性树种逐渐增多,随着环境的改变,五角槭、春榆、黄檗、白牛槭的更新优势则相对较大。改进公式与Shannon-Wiener指数计算结果基本一致,因为此时演替趋向成熟。
具有近相似环境要求的物种间生态位重叠较大。生态位重叠的结果表明,林隙内阔叶树间的种间竞争相对较强,红松与阔叶树间的竞争相对较弱。说明阔叶红松混交林比红松纯林更有利于红松种群的生长发育。
生态位宽度较大的物种对资源的利用能力较强、分布较广,因而与其它种群间的生态位重叠较大, 生态位宽度较小的物种对资源的利用能力较弱、分布不够广泛,因而与其它种群间的生态位重叠较小。具有较大生态位宽度的种群之间的生态位重叠并不一定较高。
刘建国. 1987. 生态位理论的发展及其在农村生态工程建设中的应用原则. 农业现代化研究, (6): 30-33. |
刘建国, 马世骏.1990.扩展的生态位理论.见: 马世骏主编.现代生态学透视.北京: 科学出版社
|
孙中伟, 赵士洞. 1995. 长白山北坡椴树阔叶红松林群落特征. 生态学杂志, 14(5): 26-30. |
王刚, 梁学功, 冯波. 1995. 沙漠植物的更新生态位Ⅰ.油蒿、柠条、花棒的种子萌发条件的研究. 西北植物学报, 15(5): 102-105. DOI:10.3321/j.issn:1000-4025.1995.05.019 |
王刚, 赵松龄, 张鹏云, 等. 1984. 关于生态位定义的探讨及生态位重叠计测公式改进的研究. 生态学报, 4(2): 119-127. |
臧润国, 蒋有绪, 杨彦承. 2001. 海南岛霸王岭热带山地雨林林隙更新生态位的研究. 林业科学研究, 14(1): 17-22. DOI:10.3321/j.issn:1001-1498.2001.01.003 |
张光明, 谢寿昌. 1997. 生态位概念演变与展望. 生态学杂志, 16(6): 46-51. DOI:10.3321/j.issn:1000-4890.1997.06.010 |
朱春全. 1997. 生态位态势理论与扩充假说. 生态学报, 17(3): 324-332. DOI:10.3321/j.issn:1000-0933.1997.03.017 |
Elton C S. 1927. Animal Ecology. Sidgwick and Jackson, London, 209. |
Grinnell J. 1917. The niche relationships of the California thrasher. Auk, (34): 427-433. |
Grubb P J. 1977. The maintenance of species niches in plant communities: importance of the regeneration niche. Biological Review, 52: 107-145. |
Hutchinson G E. 1957. Concluding remarks. Cold Spring Harbor Symp Quant Biol, 22: 415-427. DOI:10.1101/SQB.1957.022.01.039 |
Pianka E R.1975. Niche relations of disert Lizards.In: Cody M, Diamond J(eds.).Ecology and Evolution of Communities. Cambridge, Harvard University Press, 292-314
|