 2005, Vol. 41
2005, Vol. 41文章信息
- 郑淮兵, 董丽, 郑彩霞.
- Zheng Huaibing, Dong Li, Zheng Caixia.
- 低温和PEG“渗控”预处理促进石楠种子萌发的研究
- Improving Germination of Photinia Seed by Low Temperature and PEG Pretreatment
- 林业科学, 2005, 41(3): 54-57.
- Scientia Silvae Sinicae, 2005, 41(3): 54-57.
-
文章历史
- 收稿日期:2003-06-02
-
作者相关文章



2. 北京林业大学生物科学与技术学院 北京 100083
2. College of Biological Science and Technology, Beijing Forestry University Beijing 100083
石楠(Photinia serrulata)是蔷薇科(Rosaceae)石楠属的常绿阔叶植物,原产我国中部及南部,有较强的抗寒性,在暖温带地区有一定引种前景。目前少数成苗已在北京引种,并可在室外正常开花结实,且其结实率高,种子成熟度较好; 但石楠种子在低温下出芽率较低。大量的研究表明,PEG预处理对提高种子萌发期间的抗寒性有较好的效果(陈秉初等,1992;王飞等,1999;黄祥富等,1999)。因此本试验采用低温和不同质量分数的PEG处理种子的方法,测定其相应的生理生化指标的变化,以期为今后的引种工作提供理论依据。
1 材料与方法 1.1 材料石楠种子取自北京警察学院约19年株龄的2棵母株上。这2棵母株种植于建筑物的向阳面,在北京已有近10年的生活史,冬季能在自然条件下越冬,并有数年的开花结果史。种子采于2000年秋,采后在室外自然风干,2001年春用于试验。PEG(6 000)是日本进口分装产品。
1.2 方法 1.2.1 PEG预处理参照陈秉初等(1992)的方法,并加以改进。在21 ℃室温下,把种子分别放在不同的试管中,依次加入质量分数为1%、5%、15%、25%、35%的PEG溶液,用去离子水作对照,浸种24 h,其中每隔3 h摇动试管几次,以利于通气。每一处理设置3次重复。
1.2.2 低温胁迫处理经预处理的上述种子用蒸馏水反复冲洗,直至把药剂完全洗净,然后再把湿种子放回试管中,并立即放入0~-2 ℃的冰箱里进行低温胁迫处理24 h。
1.2.3 萌发试验经低温胁迫后,将每种处理分别取出50粒种子在发芽箱中萌发10 d,温度(21±2) ℃,光照2 000 lx, 24 h全光照。其中:萌发指数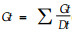
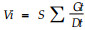
在种子进行PEG预处理的过程中,分别于处理后1、3、5、7、9 h取出10粒种子,用以测定种子的吸水曲线。
1.2.5 蛋白质含量的测定采用色素结合法(Bradford, 1976)。取植物材料0.1 g左右,加0.5 mL 50 mmol pH6.3的磷酸缓冲液研磨提取,15 000 r·min-1离心10 min后,取上清液,先测可溶性蛋白的含量,后按曾韶西等(1997)的方法测煮沸稳定蛋白含量。
1.2.6 MDA含量的测定参考赵世杰等(1994)的方法。
1.2.7 SOD活性的测定采用NBT光照化学还原法(Dhindsa, 1981)。
1.2.8 POD活性的测定采用愈伤木酚氧化法(李合生等,2001)。
1.2.9 CAT活性的测定采用高锰酸钾滴定法(李合生等,2001)。
1.2.10 蛋白质多肽的电泳分离参照Laemmli(1970)的方法。
2 结果与讨论 2.1 对种子萌发过程中的影响表 1显示了PEG预处理后,再经过低温处理对种子萌发过程的影响。从表 1可见,同对照相比,5%、15%、25%、35%的PEG预处理都能提高种子的萌发率,以15%和25%2种质量分数为最好;而15%、25%、35%3种质量分数不仅增加了幼苗的苗长(胚根和下胚轴之和),提高了幼苗的萌发指数,而且都极显著地提高了种子的活力指数。这说明15%、25%、35%3种质量分数的PEG预处理对于石楠种子来说都有较好的效果,而从综合指标上分析,15%的PEG处理效果最好。
|
|
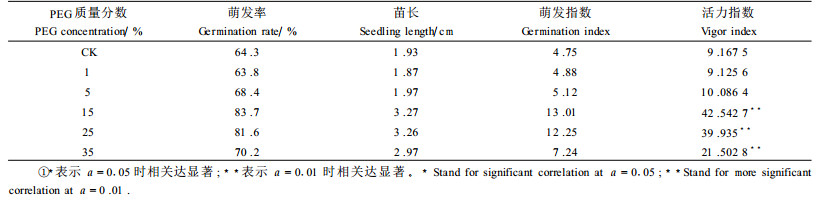
图 1的结果显示,经PEG处理后,能明显降低种子的吸水速度,而且随着PEG质量分数的升高,这种效果也随之增加。其中1%、5%2个质量分数对吸水速度的降低不太明显,25 %、35%2种质量分数能明显延缓种子吸水,15%的质量分数则处于上述两类之间。PEG处理能降低种子的吸水速度,这同郑光华等(1980)、吕小红等(1990)研究的结果相符。其原因是与它本身的特性和种子吸水时的状况有关。一般干种子细胞内含有许多大分子物质,这些物质将降低种子的衬质势,进而使种子的水势降到较低的水平。根据韩蕊莲等(1993)的研究表明:处于风干状态的种子,其水势大约为-20.0 MPa。当种子被放入水势接近于0的水中时,水分会很快地进入种子中;而在PEG溶液中,溶液的水势较低(PEG是一种高分子渗压剂,它能降低溶液的渗透势),这就相应减小了种子内部和溶液之间的水势差,进而减缓了种子的吸水速度,并且质量分数越高,这种趋势越明显。

|
图 1 PEG最初9 h里预处理对种子吸水的影响 Fig. 1 Effect of PEG pretreatment on water content in Photinia seeds during the first 9 h pretreatment |
从表 2可见,15%PEG预处理种子经低温胁迫后,其可溶性蛋白含量比对照增长了18 2%,说明经PEG预处理的种子,虽然经过一个较长时间(24 h)的低温胁迫,其可溶性蛋白的含量仍相当高,而植物受低温胁迫后可溶性蛋白含量的高低在一定程度上代表了植物细胞内各种酶以及其他大分子蛋白的含量(巫光宏等,2002)。对于刚刚萌发的种子,可溶性蛋白的含量从某种程度上反映了种子代谢活性的高低,这一结果从侧面说明低温和PEG处理能诱导萌发中种子抗寒性的提高。
|
|
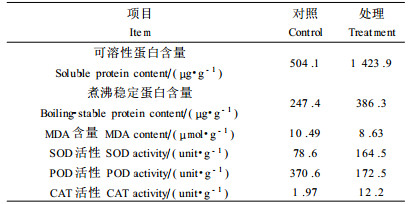
从表 2还可见,15%PEG预处理种子经低温胁迫后煮沸稳定蛋白(boiling-stable proteins)的含量比对照增高了56%。煮沸稳定蛋白一般是冷驯化、ABA或脱水等处理诱导的产物,这类蛋白一个显著的特点是具有热稳定性,即经100 ℃煮沸10 min仍留在溶液中。它们具有特异性的序列及高度的亲水性和调节渗透压的功能,推测它们有稳定膜和抗冻的作用(Lin et al., 1990;Hincha et al., 1992)。本试验的结果显示出煮沸稳定蛋白的变化趋势同可溶性蛋白的变化趋势大体一致,这同曾韶西等(1997)研究的结果相似,即水稻幼苗抗寒性的提高,伴随着可溶性蛋白及煮沸稳定蛋白的增高。
2.4 对膜脂过氧化的影响从表 2可见,PEG预处理种子经低温胁迫后,SOD、CAT活性比对照显著升高,分别提高109%和519%;MDA含量与POD活性较对照低,分别降低17.7%和53.5%。SOD的主要功能是催化超氧物阴离子自由基发生歧化反应,生成H2O2和O2,从而消除氧自由基对植物的伤害。上述结果中,经低温和PEG预处理的种子SOD活性显著升高,说明对照种子的SOD活性可能受低温胁迫影响而处于被抑制状态,而低温和PEG预处理的种子能减轻这种抑制作用。SOD虽然可清除氧自由基,但又有H2 O2的生成,H2O2主要作为氧化剂对植物组织产生毒害作用,它可以产生氧化能力很强的羟自由基和单线态分子氧,过量的H2O2积累会对植物细胞造成很大伤害。植物组织中高浓度的H2O2主要靠CAT清除,从而使H2O2控制在较低的水平(杨淑慎等,2001)。POD对植物的作用比较复杂,目前认为它是具有双重效应的一种酶。一方面为保护效应,其反应机制是POD能催化相应的基质(如酚类化合物)与H2O2反应,从而达到消除H2O2的目的。许多试验支持这一观点:戴金平等(1991)通过低温锻炼黄瓜幼苗后,发现幼苗中的POD活性增高;任小林等(1991)在研究杏果实成熟衰老过程中生理指标的变化,POD活性呈下降趋势。另一方面,POD参与叶绿素的降解、活性氧的产生,并引发膜脂过氧化,是植物衰老到一定阶段的产物,为一种伤害反应,可作为植物衰老的指标(杨淑慎等,2001)。从本试验的结果中石楠种子POD的作用推测为保护效应,其活性之所以较对照低,可能是因为此时预处理种子的CAT活性较高,可清除大量的H2O2,而POD的启动或许有一个过程,它可能需要在某种条件(如一定量的H2O2)下才得以实现。
MDA是膜脂过氧化的最终产物,已被证明MDA的积累来自不饱和脂肪酸的降解(段咏新等,1997;陆定志,1983),它是由体内自由基引发而产生的(Dhindsa et al., 1982)。因此,MDA的积累在一定程度上反映了体内自由基的活动状态,MDA积累多,说明超氧阴离子与羟自由基可能是高水平的。MDA和一些自由基过多的积累会对植物造成严重的伤害:MDA具强交联性质,能与氨基酸或有游离氨基酸的蛋白质、磷脂酰乙醇胺及核酸结合,形成具有荧光的Schiff碱,即类脂褐色素(Lipofulscin-like pigment, LFP),它是干扰细胞内正常生命活动代谢的不溶性化合物(蒋明义等,1991)。黄玉山等(1997)在研究镉诱导植物的自由基过氧化损伤时发现,MDA积累与生长量呈负相关,与质膜透性的增加呈显著正相关。
2.5 预处理种子在萌发中蛋白质多肽组成的分析图 2为PEG预处理种子经低温胁迫后,在常温萌发过程中的SDS-PAGE电泳分析图谱。从电泳图中可以看出:与对照相比,预处理的种子未引起电泳图谱谱带的增减,但在常温下萌发的第4天,在40.27、42.5、44.5、46.2、50.8kD处的5条蛋白带的染色比未处理的深得多。梁峥等(1989)在用PEG引发大豆种子时,通过SDS-PAGE电泳图谱分析,发现引发种子和未引发种子的可溶性蛋白带的数目并没有变化,但引发种子蛋白带的染色比未引发种子的蛋白带染色深。本试验得到类似的结果,这说明PEG预处理后的种子,经低温胁迫后,部分多肽的含量高于对照水平,这些多肽含量的增加与可溶性蛋白含量及部分酶活性的增加有关,它是否还导致了其他抗寒性物质的增加,还需要作进一步研究。

|
图 2 PEG预处理种子在萌发中的SDS-PAGE电泳图谱 Fig. 2 Examination of polypeptides of PEG-pretreated seeds extracted by SDS-PAGE during 5 days of germination 1.处理种子低温胁迫后;2.对照种子低温胁迫后;3.处理种子萌发第2天;4.对照种子萌发第2天;5.处理种子萌发第3天;6.对照种子萌发第3天;7.处理种子萌发第4天;8.对照种子萌发第4天;9.标准蛋白;10.处理种子萌发第5天。 1. Pretreated seeds immediately treated with cold stress; 2. Control immediately treated with cold stress; 3. After 2 days' germination of treated seeds; 4. After 2 days' germination of control seeds; 5. After 3 days' germination of treated seeds; 6. After 3 days' germination of control seeds; 7. After 4 days' germination of treated seeds; 8. After 4 days' germination of control seeds; 9. Mark; 10. After 5 days' germination of treated seeds. |
陈秉初, 吴美仙, 汪久康. 1992. PEG预处理对早稻种子活力和抗寒能力的影响. 植物生理学通讯, 28(3): 198-201. |
戴金平, 沈征言, 简令成. 1991. 低温锻炼对黄瓜幼苗几种酶活性的影响. 植物学报, 38(8): 627-632. |
段咏新, 李松泉, 傅家瑞, 等. 1997. 钙对延缓杂交水稻叶片衰老的作用机理. 杂交水稻, 12(6): 23-25. |
韩蕊莲, 侯庆春, 邹厚远, 等. 1993. 用PEG"引发"沙打旺种子活力及抗逆性的研究. 草业科学, 10(6): 60-63. |
黄祥富, 蒋明兰, 廖军. 1999. PEG渗调对苦瓜种子活力和膜脂过氧化的影响. 种子, (2): 7-9. |
黄玉山, 罗广华. 1997. 镉诱导植物的自由基过氧化损伤. 植物学报, 39(6): 522-526. |
李合生, 孙群, 赵世杰, 等. 2001. 植物生理生化实验原理和技术. 北京: 高等教育出版社.
|
梁峥, 珞爱玲, 邹喻苹, 等. 1989. 聚乙二醇引发对冷敏感大豆种子蛋白质合成的影响. 科学通报, 34(11): 862-864. |
陆定志. 1983. 叶片的衰老及其调节控制. 植物生理生化进展, (2): 20-52. |
蒋明义, 荆家海, 王韶唐. 1991. 渗透胁迫对水稻幼苗膜脂过氧化及体内保护系统的影响. 植物生理学报, 17(1): 80-84. DOI:10.3321/j.issn:1671-3877.1991.01.014 |
吕小红, 傅家瑞. 1990. 聚乙二醇渗调处理提高花生种子活力和抗寒性. 中山大学学报(自然科学版), 29(1): 63-70. DOI:10.3321/j.issn:0529-6579.1990.01.003 |
任小林, 李嘉瑞. 1991. 杏果实成熟衰老过程中活性氧和几种生理指标的变化. 植物生理学通讯, 27(1): 34-36. |
王飞, 丁勤, 杨峰. 1999. PEG预处理对老化杜梨种子活力的影响. 种子, (4): 20-22. |
巫光宏, 詹福建, 罗焕亮. 2002. 几种保护酶活性变化与马占相思树对低温胁迫的抵抗性的关系研究. 植物研究, 22(1): 42-45. DOI:10.3969/j.issn.1673-5102.2002.01.013 |
杨淑慎, 高俊凤. 2001. 活性氧、自由基与植物的衰老. 西北植物学报, 21(2): 215-220. |
曾韶西, 王以柔, 李美如. 1997. 不同胁迫预处理提高水稻幼苗抗寒性期间蛋白质的变化. 植物学报, 39(2): 130-136. |
赵世杰, 许长成, 邹琦, 等. 1994. 植物组织中丙二醛测定方法的改进. 植物生理学通讯, 30(3): 207-210. |
郑光华, 顾增辉, 徐本美. 1980. 瓜尔豆种子发芽生理的研究. 植物生理学报, 6(2): 115-125. |
Bradford M M. 1976. A rapid and sensitive method for the quantification of microgram quantities of proteins utilizing the principle of protein-dry binding. Anal Biochem, 72: 248-254. DOI:10.1016/0003-2697(76)90527-3 |
Dhindsa R S, Plumb-Dhindsa P L, Reid D M. 1982. Leaf senescence and lipid peroxidation: effect of some phythohormones, and scavengers of free radicals and singlet oxygen. Plant Physiol, 56: 453-457. DOI:10.1111/j.1399-3054.1982.tb04539.x |
Dhindsa R S. 1981. Leaf senescence correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoid dismutase and catalase. J Exp Bot, 32: 93-101. DOI:10.1093/jxb/32.1.93 |
Hincha D K, Schmitt J M. 1992. Cryoprotective leaf proteins: assay methods and heat stability. Plant Physiol, 40: 236-238. |
Laemmli U K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685. DOI:10.1038/227680a0 |
Lin C, Guu W W, Everson E, et al. 1990. Cold acclimation in Arabidopsis and wheat: a response associated with expression of related gens encoding `Boiling-stable' polypeptides. Plant Physiol, 94: 1078-1081. DOI:10.1104/pp.94.3.1078 |