 2005, Vol. 41
2005, Vol. 41文章信息
- 赵凤君, 高荣孚, 沈应柏, 苏晓华, 张冰玉.
- Zhao Fengjun, Gao Rongfu, Shen Yingbai, Su Xiaohua, Zhang Bingyu.
- 水分胁迫下美洲黑杨不同无性系间叶片δ13C和水分利用效率的研究
- A study on foliar carbon isotope composition(δ13C) and water use efficiency of different Populus deltoides clones under water stress
- 林业科学, 2005, 41(1): 36-41.
- Scientia Silvae Sinicae, 2005, 41(1): 36-41.
-
文章历史
- 收稿日期:2004-04-02
-
作者相关文章



2. 中国林业科学研究院林业研究所 北京 100091
2. The Research Institute of Forestry, CAF Beijing 100091
美洲黑杨(Populus deltoides)是当今世界中纬度地区最适合的短轮伐期工业用材集约经营树种之一,近年来我国引进了许多优良的美洲黑杨品系,在全国范围内推广已获得很好的经济效益。美洲黑杨速生,但不可否认,其耗水量很大。在世界性的水资源亏缺面前,快速、高效地选育速生高水分利用效率(WUE)的无性系已成为迫切需求,而完成此项工作的关键是准确地测定WUE,但这也是长期以来困扰育种工作者的难题。WUE的直接测定方法有2种:个体或群体水平的WUE也称WUEL,等于植物在较长期生长过程中形成的干物质量和耗水量之比,一般以每克水产生多少毫克干物质来表示。此方法准确,符合实际需要,但费时费力。单叶水平的WUE也称瞬时WUE,是用瞬时测定的净光合速率(Pn)和蒸腾速率(Evap)之比来表示,单位是μmolCO2·mmol-1H2O。瞬时WUE测定简单,但只代表某特定时间内植物部分叶片的行为,和WUEL没有很好的相关性(Martin, 1988)。现在国际常用的一种间接测定WUE的方法是稳定碳同位素技术,Farquhar等(1982)在大量试验数据积累和相关理论推导的基础上,认为从理论上稳定碳同位素分辨力(Δ)和WUE呈负相关,即稳定碳同位素组成(δ13C)和WU E正相关,并首先在小麦(Triticum aestivum)上得到证实。稳定碳同位素技术通过对长期积累于叶片或其他器官中的碳代谢产物的稳定碳同位素分析来评估叶片或植株生长过程中总的WUE特性,比瞬时WUE更具代表性,且其测定简单、不受时间和季节的限制,样品采集烘干之后,其中的碳同位素成分就不再改变(林植芳等,2001)。各国科研工作者在小麦(Malse et al., 1988)、花生(Arachis hypogaea)(Hubick et al., 1988)、棉花(Gossypium hirsutum)(Hubick et al., 1987)、大麦(Hordeum vulgare)(Hubick et al., 1989)等农作物上进行了大量研究,大部分试验结果显示Δ和WUE负相关;但也有Δ和WUE正相关和根本不相关的报导(Farquhar et al., 1989)。近年来,国外对用于人工林集约经营的树种其不同基因型的Δ和WUE的关系有较广泛的研究。Sun等(1996)对白云杉(Picea glauca),Osorio等(1994)对蓝桉(Eucalyptus globulus)的研究都认为,在各种水分条件下高δ13C都可作为高WUE无性系的可靠指标。Lawrence等(199 5)等对黑云杉(Picea mariana)的研究则认为,只有在干旱条件下Δ和WUE负相关,其他条件下没有相关性。国内对δ13C的研究起步较晚,对于不同基因型Δ和WUE的关系的研究集中在农作物方面。林植芳等(2001)对大豆和小麦,薛慧勤等(1999)对花生的研究都认为,低Δ可作为筛选高WUE的可靠指标。而林木在此方面的研究集中在不同树种及同一树种在不同条件下的δ13C差异性上,严昌荣等(1998;2002)、韩兴国等(2000)认为δ 13C在种间差异很大,且各种环境因子都可引起δ13C的变化,同一树种在土壤干旱条件下WUE会升高(严昌荣等,2001)。本文对美洲黑杨不同无性系叶片δ13C、水分胁迫下的生物量、WUEL的相关性进行了初步研究,以期为高WUE的美洲黑杨无性系选育提供理论依据和方法。
1 材料和方法 1.1 试验材料试验材料为美洲黑杨杂交无性系,其父本为哈佛杨(P. deltoides cv. ‘Harvard'),母本为山海关杨(P. deltoides cv. ‘Shanhaiguanensis'),共10个无性系,编号为J1,J2…J10。当前优良品系108杨(P. euramericana cv. ‘114/69')和中林46杨(P. deltoides cv. ‘Zhonglin 46')为对照,编号为CK1、CK2。
1.2 试验处理试验在中国林业科学研究院温室内进行,温室顶部有天窗,下雨时关闭,其他时间全部打开,保证空气畅通;温室全部由玻璃制成,透光良好;温室内无特殊的控温、控湿措施,6—9月份8:00平均温度为26 ℃,平均相对湿度为75%;13:00平均温度为33 ℃,平均湿度为5 5%;极端最高温为40 ℃,出现在8月份;温室内CO2浓度变化不大,在0.38 μmol·mol -1左右。
无性系插穗于2003年4月1日栽于塑料盆中,盆高30 cm, 内径25 cm。培养土为苗圃熟土(10份)、细沙(2份)、草炭土(1份)混合而成,田间持水量为20.20%, 土壤容重为1.274 g·cm-3。各无性系插穗大小和质量尽可能保持一致,每盆栽1株,在充分供水条件下培养。6月10日苗高达30 cm左右时,控水试验开始,至8月20日结束。控水试验采用完全随机区组设计,共有4个水分处理:充分供水、轻度胁迫、中度胁迫、严重胁迫,土壤含水量分别控制在田间持水量的70%~90%、60%~80%、50%~70%、40%~60%。同一水分处理下所有苗木浇水量相同,各处理的浇水量累计值分别是16.5 L、11.5 L、8.2 L、6.7 L。每处理每品系有9株苗。用烘干和称重相结合的方法控制土壤含水量,每半个月施肥一次,充分保证养分的供应。
1.3 测定项目与方法 1.3.1 生物量生物量共测定2次,分别在6月10日、8月20日,控水试验开始前和结束后。在苗高、地径测定的基础上,各品系选生长中等的苗木4株,拔出、洗净,放入烘箱中105 ℃杀青1/2 h,然后70 ℃烘至恒重,约72 h。用1/10 000电子感应天平称干物质量,取均值为最后结果,用减重法测得控水期间生物量的累计值(单位:g)。
1.3.2 长期水分利用效率(WUEL)控水期间生物量(干物质量)与累计浇水量之比,单位为mg·mL-1。
1.3.3 稳定碳同位素组成(δ13C)在控水试验结束时,各处理、各品系选生长中等的苗木3株,每株取1片叶(第9片叶),共取3叶片混合后放入烘箱中,首先105 ℃杀青1/2 h,然后70 ℃烘至恒重(约72 h),研磨过筛,经高温燃烧成CO 2,用MAT-251型质谱仪测定样品的13C/12C,并据δ13C=(Rp-Rs)/Rs (Farquhar et al., 1989)分别计算δ13C(在中国农业科学院原子能研究所完成)。式中:Rp、Rs分别表示植物组织样品和标准化石PDB的13C/12C。
2 结果与分析 2.1 控水期间的生物量由图 1可看出,随干旱胁迫的加剧,各无性系生物量都呈现明显下降趋势。J2在4个水分处理中,生物量都是最优的,是适应面较宽的一个优良无性系。表现较优的还有J7、J8、J9,逆境下的生物量都显著超过对照。J6在充分供水和轻度胁迫下生物量表现不突出,分别排在第7、10位,但中度胁迫和严重胁迫下则排在仅次于J2的第2位,说明J6是一个适合于栽植在干旱逆境下的优良无性系。

|
图 1 不同水分处理下各无性系控水期间的生物量 Fig. 1 Total biomass of different clones under different water treatments |
充分供水、轻度胁迫、中度胁迫、严重胁迫下无性系间WUEL的单因素方差分析结果都显示差异显著,进一步的多重比较结果(表 1)显示:充分供水条件下,除J1、J5、J10外其他无性系间WUEL差异不显著;轻度胁迫条件下,无性系间WUEL差异显著,且分为几个等级,J2最优,其次为J7、J9、J3、J8、CK1,再次为CK2、J1、J4、J6,最差为J5、J10;中度胁迫条件下,J2最优,其次为J6、J9、J7,再次为J8,最差为J5、J10;严重胁迫条件下,最优仍为J2,其次为J6、J9、J8、J7,再次为J4、J3、J10,最差为J5。以上分析表明,无性系间WUEL的差异随水分胁迫的加剧而更加显著,J2在各水分条件下WUEL都是最优的,其次为J6、J7、J8、J9。须特别指出的是J6,此无性系在充分供水和轻度胁迫条件下,WUEL不突出,但在中度胁迫和严重胁迫下它的WUEL仅次于J2,这说明J6是一个适应干旱逆境的优良无性系。
|
|
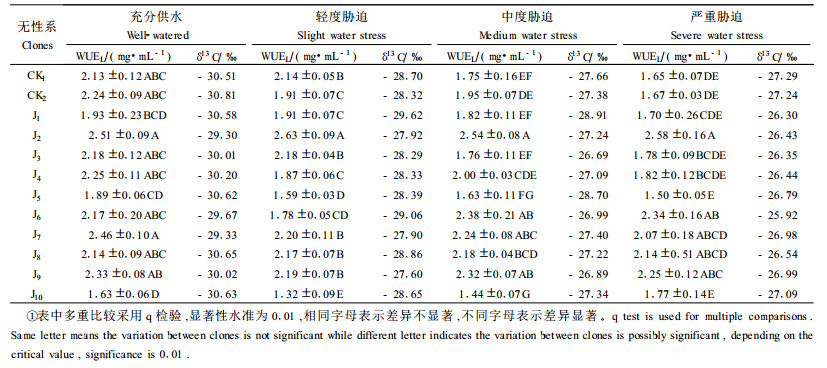
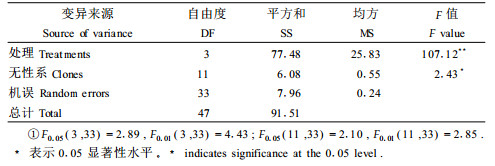
由表 1数据可看出,各无性系叶片δ13C随水分胁迫的加剧呈明显上升趋势,严重胁迫比充分供水平均上升了3.50‰;同一水分处理条件下无性系间δ13C也存在差异,极差分别为1.51‰、2.02‰、2.22‰、1.37‰,均值为1.78‰。
由表 2的双因素方差分析结果可看出,δ13C在水分处理间达到极显著水平,在无性系间也达到显著水平;通过对均方(25.83和0.55)和F(107.12和2.43)值的比较分析,可看到水分处理引起的δ13C差异要远大于无性系间的差异,因此水分是引起δ13C的差异的主要因素。
|
|
由以上分析可知,同一水分条件下无性系间δ13C和WUEL差异都很显著。图 2的曲线显示,CK1、CK2、J2、J6、J7、J8、J9的δ13C和WUEL在各水分处理条件下都呈现正相关,相关系数分别为0.7491、0.7019、0.7327、0.77 56。

|
图 2 δ13C和WUEL的相关性分析 Fig. 2 The relationship between δ13C and WUEL a.充分供水Well-watered(y=3.065 2x+129.78, r=0.749 1);b.轻度胁迫Slight water stress(y=4.2098 x+143.47, r=0.7019);c.中度胁迫Medium water stress(y=6.192 2x+186.77, r=0.732 7);d.严重胁迫Severe water stress(y=3.578 1x+0.601 6, r=0.775 6). |
同等水分条件下美洲黑杨杂交无性系间WUEL差异极显著,J2、J6、J7、J8、J9是WUEL显著高于对照及其他的优良无性系。表 1数据显示,J2、J6、J7、J8、J9的WUEL在水分胁迫下变化不大,保持在2 m g·mL-1以上,而对照及其他无性系WUEL呈明显下降趋势。武维华(2003)认为干旱胁迫下植物的WUEL会升高,且Dickmann等(2002)认为黑杨派杨树每消耗1 mL水可积累3.5~4.4 mg干物质,可表 1中的WUEL偏低,最大为2.63 mg·mL-1,出现这种情况是因为本文以总浇水量作为计算WUEL的基数,没有去除土壤蒸发所消耗的水量。水分胁迫下的土壤蒸发量占浇水量的比例要远大于充分供水条件下的比例,因此若仅以植株蒸腾量为基数计算WUEL,则胁迫条件下的值要大于表 1中的值,即WUEL会呈现上升趋势,这和Farquhar等(1982)及大部分研究人员的结论(William et al., 2003; Miguel et al., 2000; Zhang et al., 1993; 严昌荣等,2001)是一致的。本文之所以要采取同一处理所有苗木浇水量都相同的处理方式,是因为此种方法更接近于大田条件下的土壤自然干旱过程,且这种方法更适合于人工林的集约经营管理,可以很轻松地比较出同等浇水量条件下各无性系总生物量的差异。
3.2 δ13C和WUEL的相关性同等水分条件下,δ13C和WUEL正相关,δ13C是间接评估美洲黑杨杂交无性系间WUEL差异的可靠指标。自然条件下有2种稳定性碳同位素,其中12C占98.89%,13C占1.108%。将植物组织样品中的13C/12C与产自美国南卡罗纳州的标准化石样品PDB〔 Pee Dee Belemnite, 一种海洋中的贝壳化石,其13C含量为1.124%(Farquhar et al., 1989)〕中的13C/12C相比较,偏离值以δ1 3C表示。不同植物或同种植物在不同环境条件下其δ13C差异很大。Farquhar等(1982)把植物对13C的分辨力定义为Δ,δ13C越大Δ越小;Δ由气孔扩散分辨力和羧化酶分辨力的差异而引起,对于C3植物,碳代谢过程中只有一种Rubisco酶,因此同一种C3植物δ13C变化的主要影响因子是气孔。气孔的行为特征既受遗传因子决定,同时又对环境因子非常敏感,其中最重要的因子是水分。对于C3植物,同等水分条件下无性系间由于气孔行为的差异,如气孔密度、大小、不均匀开闭程度等会导致气孔导度GS的差异,GS较低的品系蒸腾速率Evap小,胞间CO2浓度(Ci)/Ca低。GS对净光合速率Pn的影响要小于对Evap的影响,因此尽管无性系间Ci/Ca存在差异,但无性系间Pn差异不会很大,在这种情况下Ci/Ca较低的无性系WUE就会较高,而同时因其有较多的13C被利用,而导致δ13C较高,这就是高δ13C指示高WUE基因型的理论基础。
同等水分条件下基因型间Ci/Ca的差异是由内在的遗传因素决定的,是WUE差异的基础。环境中的胁迫因子特别是水分,可引起GS降低,进而引起Ci/C a降低,且降低程度远超出基因型间的差异。表 1水分胁迫导致δ13C变化值为3.50‰,远大于同一水分条件下品系间的差异1.78‰。表 2的双因素方差分析也说明了这一点。因此在对无性系间的δ13C值进行比较时,必须严格限制水分条件,不同水分条件下的δ13C不能进行对比。许多研究结果显示,δ13C和WUE之间负相关或根本不相关(Farquhar et al., 1989),可能就是由于没有严格控制水分条件。另外,当苗木在干旱胁迫下生长受到抑制时,也可造成Ci/Ca降低,从而引起δ13C升高。从表 1看,中等胁迫下优良品系J2和WUEL较小品系J3的δ13C值分别为-27.24‰,-26.69‰,严重胁迫下J2和WUEL较小品系J1的δ13C值分别为-26.4 3‰,-25.8‰。以上2因素导致水分胁迫下各品系δ13C和生物量的相关性降低,这也是本文中δ13C和WUEL相关系数仅有0.7的原因。
韩兴国, 严昌荣, 陈灵芝, 等. 2000. 暖温带地区几种木本植物碳稳定同位素的特点. 应用生态学报, 11(4): 497-500. DOI:10.3321/j.issn:1001-9332.2000.04.005 |
林植芳, 彭长连, 林桂珠. 2001. 大豆和小麦不同基因型的碳同位素分馏作用及水分利用效率. 作物学报, 27(4): 410-414. |
武维华. 2003. 植物生理学. 第1版. 北京: 科学出版社, 444-446.
|
薛慧勤, 甘信民, 孙明辉, 等. 1999. 干旱条件下花生水分利用效率与叶片碳同位素辨别力的相关性研究. 中国油料作物学报, 21(1): 27-34. DOI:10.3321/j.issn:1007-9084.1999.01.009 |
严昌荣, 韩兴国, 陈灵芝. 2001. 六种木本植物水分利用效率和其小生境关系研究. 生态学报, 21(11): 1952-1956. DOI:10.3321/j.issn:1000-0933.2001.11.028 |
严昌荣, 韩兴国, 陈灵芝, 等. 1998. 暖温带落叶阔叶林主要植物叶片中δ13C值的种间差异及时空变化. 植物学报, 40(9): 853-859. DOI:10.3321/j.issn:1672-9072.1998.09.014 |
严昌荣, 韩兴国, 陈灵芝, 等. 2002. 中国暖温带落叶阔叶林中某些树种的13C自然丰度:δ13C值及生态学意义. 生态学报, 22(12): 2163-2166. DOI:10.3321/j.issn:1000-0933.2002.12.021 |
Dickmann D I, Isebrands J G, Blake T J, et al.2002. Poplar culture in North America.Ottawa, Canada: NRC Research Press, National Research Council of Canada, 77-118
|
Farquhar G D, Ehleringer J R, Hubick K T. 1989. Carbon isotope discrimination and photosynthesis. Annu Rev Plant Physiol Plant Mol Biol, 40: 503-537. DOI:10.1146/annurev.pp.40.060189.002443 |
Farquhar G D, O'Leary M H, Berry J A. 1982. On the Relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust J Plant Physiol, 9: 121-37. |
Farquhar G D, Richards R A. 1984. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Aust J Plant Physiol, 11: 539-552. |
Hubick K T, Farquhar G D. 1987. Carbon isotope discrimination selecting for water use efficiency. Aust Cotton Grower, 8: 66-68. |
Hubick K T, Farquhar G D. 1989. Genetic variation of transpiration efficiency among barley genotypes is negatively correlated with carbon isotope discrimination. Plant Cell Environ, 24: 92-99. |
Hubick K T, Shorter R, Farquhar G D. 1988. Heritability and genotype environmental interaction of carbon isotope discrimination and transpiration efficiency in peanut(Arachis hypogaea). Australian Journal of Plant Physiology, 15: 799-813. |
Lawrence B F, Kurt H J. 1995. Genetic variation in carbon isotope discrimination and its relationship to growth under field conditions in full-sib families of Picea mariana. Can J For Res, 25: 39-47. DOI:10.1139/x95-005 |
Malse J, Farquhar G D. 1988. Effects of soil strength on the relation of water-use efficiency and growth to carbon isotope discrimination in wheat seedlings. Plant Physiol, 86: 32-38. DOI:10.1104/pp.86.1.32 |
Martin B, Thorstenson Y R. 1988. Stable carbon isotope composition(δ13C), water-use efficiency, and biomass productivity of Lycoperscon esculentum, Lycoperscon pennellii, and the F1 hybrid. Plant Physiol, 88: 213-217. DOI:10.1104/pp.88.1.213 |
Miguel O G, Bert M C, Thomas C H. 2000. Genotypic variation in carbon isotope discrimination and gas exchange of ponderosa pine seedlings under two levels of water stress. Can J Res, 30: 1581-1590. DOI:10.1139/x00-080 |
Osorio J, Pereira J S. 1994. Genotypic differences in water use efficiency and 13C discrimination in Eucalyptus globulus. Tree Physiology, 14: 871-882. DOI:10.1093/treephys/14.7-8-9.871 |
Sun Z J, Livingston N J, Guy R D, et al. 1996. Stable carbon isotopes as indicators of increased water use efficiency and productivity in white spruce(Picea glauca(Moench) Voss) seedlings. Plant, Cell and Environment, 19: 887-894. DOI:10.1111/j.1365-3040.1996.tb00425.x |
William L B, Whitlow T H, Setter T L, et al. 2003. Ecophysiology of Acer rubrum seedlings from contrasting hydrologic habitats: growth, gas exchange, tissue water relations, abscisic acid and carbon isotope discrimination. Tree Physiology, 23: 841-850. DOI:10.1093/treephys/23.12.841 |
Zhang J W, John D M. 1993. Population differences in water-use efficiency of well-watered and water-stressed western larch seedlings. Can J For Res, 24: 92-99. |