 2005, Vol. 41
2005, Vol. 41文章信息
- 杨大荣, 彭艳琼, 赵庭周, 王秋艳, 徐磊.
- Yang Darong, Peng Yanqiong, Zhao Tingzhou, Wang Qiuyan, Xu Lei.
- 聚果榕果内种子季节变化与聚果榕小蜂活动的关系
- Relationship between Activity of Ceratosolen fusciceps and Seasonal Changes of the Seed Number of Ficus racemosa
- 林业科学, 2005, 41(1): 25-29.
- Scientia Silvae Sinicae, 2005, 41(1): 25-29.
-
文章历史
- 收稿日期:2003-04-11
-
作者相关文章


榕树是桑科(Moraceae)榕属(Ficus)中的乔木树种的总称。榕树及其传粉昆虫作为协同进化的典型代表,得到了广泛而深入的研究(Ramirez, 1970; Janzen, 1979; Herre, 1989; Weiblen, 2002)。全世界的榕树约有750种(Janzen, 1979; Berg, 1989),每一种榕树专一性地由一种小蜂传粉,极少例外;每一种榕小蜂只在其唯一的宿主榕树内生长繁殖,二者的互惠共生关系是协同进化系统中最为特化的一类,它们之间的关系已发展到一对一、不能互缺的高级阶段(Wiebes, 1979; 马炜梁等,1997)。榕树没有榕小蜂传粉就不会受精形成种子,没有种子就无法进行有性繁殖,失去有性繁殖功能,种群就不会自然地发展起来,种群只会逐步萎缩直至消亡;榕小蜂没有榕树就失去生活和繁殖场所,导致榕小蜂种类和群落的逐步消亡。榕属植物主要分布在热带、南亚热带地区,它是热带雨林中的最大的木本属种之一(吴征镒,1995)。榕树是所有树种中结实量最高的,在热带地区,一年四季均会结果,年结实高达3~8次,是任何一种植物都难以比拟的,在其他植物挂果量稀少的冬春季,它为多种鸟类、蝙蝠、猴子、松鼠、昆虫提供食物,是热带森林生态系统中的关键类群(Janzen, 1979;许再富,1994;杨大荣等,2000)。榕果由花序托发育而成, 肉质球形或梨形, 内着生花朵,是产生榕树种子和榕小蜂发育的场所。雌雄同株的榕树,果壁内着生雄花、瘿花和雌花;雌雄异株的榕树,雄株榕果内着生雄花和瘿花,雌株榕果内只着生雌花。雌花产生种子,瘿花内寄生榕树的传粉昆虫榕小蜂和其他非传粉小蜂。榕果的发育分为5个时期,即雌花前期(或称:幼花期)、雌花期(雌花开放期)、间花期(受精子房发育或雄花生长期)、雄花期(花粉成熟)和果实成熟期(Galil et al., 1968;彭艳琼等,2003;王秋艳等,2003)。
聚果榕(Ficus racemosa)是亚洲热带雨林中一个广布种,也是西双版纳热带雨林中的一个优势种群(West et al., 1994; 吴征镒,1995; 杨大荣等, 2000; 2001;2002a;2002b)。在西双版纳热带雨林中,聚果榕主要生长在原始雨林内部低洼处,尤其是在沟谷、河、江边,常以5~10株为一组群、株间距离50~120 m地集团性分布,很少有例外(徐磊等,2003;杨大荣等,2000; 2001;2003)。本研究连续3年,取不同月份的聚果榕的榕果,统计榕果内瘿花、种子、败育花(长花柱雌花和短花柱雌花)的数量,并进行了定量放蜂试验。分析榕果内小花的年周期变化特征,以揭示榕树与榕小蜂的共生机制。
1 材料与方法 1.1 研究样地样地设在西双版纳州勐腊县勐仑镇,海拔560~580 m,年均气温21.4 ℃,最热月是4月,月均气温33.5 ℃, 最冷月为1月, 月均气温11.2 ℃,年降雨量1 557.0 mm, 相对湿度86%。全年四季不分明,但有明显的干湿季节交替,每年的11月至翌年的4月为干季(雾凉季11月— 2月和干热季3—4月),降雨量仅占全年的13%~17%,5—10月为雨季,降雨量占全年的82%~87%。根据地理位置、生态环境、人为干扰程度等因素选择2个研究样地。
1号样地:中国科学院西双版纳热带植物园罗梭江边,位于植物园吊桥西面的竹林,海拔560 m。西、北为罗梭江,东南面是竹林,正东为柚子园。为砍伐残存的热带原始林和次生林地,植被覆盖度为75%,人为干扰严重。样地内有聚果榕13株。
2号样地:云南省西双版纳州勐仑自然保护区,位于小腊公路56 km处,海拔570~580 m。系1958年建立的勐仑自然保护区主体,样地内热带季雨林保护完整,植被覆盖度为90%~95%,人为干扰较少。样地内有聚果榕9株。
1.2 研究材料聚果榕:高大乔木,高25~30 m,胸径60~240 cm。聚果榕开花具有典型的树内同步、树间不同步的特征。同一株树内的榕果同时开花,同时成熟;不同榕树之间的花期相互错开,一年四季都有榕树开花,为榕小蜂的生活和繁殖提供了保障。聚果榕是雌雄同株同果植物,榕果内同时生有长花柱雌花、短花柱雌花和雄花。长花柱雌花经榕小蜂传粉受精后发育成种子;短花柱雌花则供给榕小蜂繁殖性雌蜂产卵繁殖后代。同一榕果内的雌花和雄花成熟期异步,雌花开放是在榕果发育8~15 d的雌花期,该时期雄花尚在发育;当榕果生长发育到32~45 d,种子和瘿花内小蜂发育成熟时,雄花方成熟,花药裂开,释放出花粉。榕树之间传粉成功与否与榕小蜂寿命和种群大小相关,榕小蜂种群大小直接受榕树种群大小的影响;由于只有雌性榕小蜂才能授粉,因此,传粉也和榕小蜂的性比有关。种子和瘿花产生的数量将反映榕小蜂的授粉情况。榕果内瘿花的变化可反映小蜂种群变化情况。榕果成熟时橙红色。在西双版纳热带雨林中,聚果榕一年产成熟果实4~6次。
1.3 研究方法 1.3.1 榕果内种子、瘿花、败育花的材料取样与统计在西双版纳,聚果榕果的发育从幼花期到果实成熟期,干热季和雨季需要38~46 d,而雾凉季则需要48~61 d。样本取雄花期的榕果,该时期种子和瘿花已发育完全,榕小蜂开始羽化,容易区分榕果内的各种小花。从2000年1月至2002年12月期间,隔月取样一次,每月下旬取样,每次取雄花期发育的榕果150个,随机挑取50个,测量果径,解剖分类统计榕果内瘿花、种子、败育雌花的数量,共统计2 450个榕果。统计分析比较各月份间的差异。
1.3.2 聚果榕小蜂传粉效率的试验用120筛目的绢纱网缝制成10 cm×15 cm的小口袋,在聚果榕榕果发育到幼花期时套住单果隔离,当榕果发育到雌花期,腔内小花开放、花托口松动时,捕捉在雌花期果外飞舞的小蜂。按1、2、4、6、8、10、15、30只组合放入隔离袋内,让其自然钻进果腔,每天检查3次进蜂情况,有死蜂及时补加,直至进蜂数符合设计要求止。记录进蜂数量,验证单蜂或不同组合的小蜂传粉效率和产卵繁殖效率,每个组合5个重复。当聚果榕成熟时,让榕小蜂自然出蜂到袋子后,开始收蜂,剖开果实统计每个试验果的种子量、瘿花量和败育花量。统计分析比较不同进蜂数对种子数、瘿花数、败育花的影响。
1.3.3 对非传粉小蜂的统计在每一样地采集接近成熟的榕果,每次采集样本不少于30个,全年总计每个样地采集样果不少于180个。室内用120筛目(20 cm ×15 cm)的绢纱袋单果分装,并将样果掰开以辅助羽化小蜂出果,单果隔离系紧袋口,待瘿花内所有小蜂羽化完成时,用棉球蘸乙酸乙酯将小蜂熏死,以防飞走,然后完整地收集每个果内羽化出来的小蜂,置于75%酒精瓶内保存,单果小蜂单瓶放置,并做好采集记录和标签。小蜂出完后,对榕果内种子和败育花进行记数和统计比较。对收集的各类小蜂标本,在实验室内,借助Olympu s(SZX12)体视显微镜对各类小蜂进行分类、形态拍照,统计各单果内各种小蜂的数量。
2 结果与分析 2.1 不同季节种子产生比率的变化规律植物能否产生大量种子让种群繁衍,是植物生存发展能力的重要指标。榕树产生种子,也是为了产生更多的后代,繁衍种群。聚果榕一年四季均开花结果,不过,在不同月份,种子产生比率却有显著差异(表 1)。
|
|
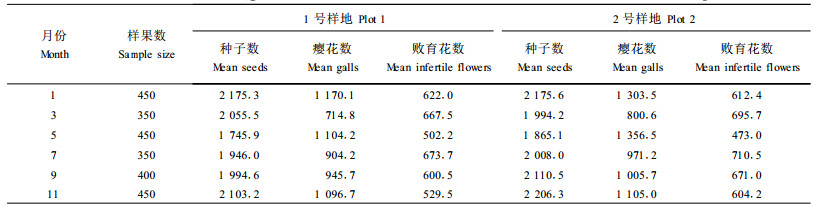
聚果榕产生种子最多的月份是1月份,该时期是森林中其他树种果实最稀少的时节,其次是1 1月份和3月份,也就是说,榕树在旱季产生的种子比雨季多。西双版纳的气候分为明显的旱季和雨季。雨季,森林中食物丰富,动物可选择的食物种类也较多。到了旱季,也就是冬春季节,森林中其他树种很少挂果,是一年中食物缺少的季节。聚果榕在这个季节产生较多的种子,适应于让更多的动物取食和传播种子,是对生境的高度适应。因为在旱季,食物稀少,使一些原来不喜欢取食榕果的动物为了生存,也以榕果为食。种子产生最少的月份是5月份。这时,一年中的高温天气已过去,刚进入雨季,是产生榕果高峰期的前夕,榕树贡献较多的能量给榕小蜂繁殖后代,扩大榕小蜂种群,为产生更多的种子做准备。
在2个样地中,总体来说,生态环境好的2号样地的种子数量要比1号样地多。但是在雾凉季的1—3月份则比1号样地的种子数低,说明由于原始林地森林密度大,阳光照射差,雾凉季较低温恒定,不利于种子发育。
2.2 聚果榕小蜂进榕果内数量对榕树种子形成的影响聚果榕的榕果成团生于老茎的瘤状短枝上,梨形,直径约2.4~3.6 cm, 果实在榕属中属中等偏大,每个榕果内有雄花56~148朵,有雌花3 584~6 520朵,平均每果4 764朵。在这些雌花之中,长柱花占总雌花数的68%,短柱花占32%。膜翅目(Hymenoptera)榕小蜂科(Agaonidae)聚果榕小蜂(Ceratosolen fusciceps)是聚果榕唯一的传粉昆虫。长柱花被聚果榕小蜂传粉受精后形成种子,短柱花提供给聚果榕小蜂或其他非传粉小蜂产卵繁殖后代。在不同季节、不同的榕果内聚果榕小蜂数量不尽相同,所以有效授粉数量不同,因此对种子形成率也不相同。解剖雌花期自然进蜂的7 840个榕果,检查果内传粉榕小蜂数量,每果进蜂1~41头不等,平均每果5.8头。经套袋定量放蜂观察,不同进蜂数量对种子形成结果均不同(图 1, 表 2)。

|
图 1 传粉小蜂数量与种子和瘿花数量的关系 Fig. 1 The relationship between the number of pollinators and the number of seeds and galls |
|
|
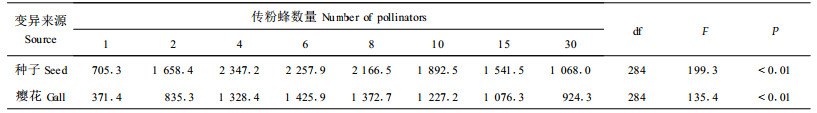
从图 1和表 2看出,聚果榕小蜂进果内传粉蜂数在4~8头最佳,特别是每个果内进去4~6头时,雌花受精形成种子率和榕小蜂繁殖率均是最高的。当每果进聚果榕小蜂低于2头时,它们身上携带的花粉量不足给长柱花传粉,而且虫体内怀的虫卵也不能在大部分短柱花上产卵繁殖,浪费了聚果榕果内的小花资源。当每个榕果内进聚果榕小蜂超过10头以上时,小花资源又不够小蜂传粉和产卵,由于蜂多干扰大,致使许多小蜂不能正常传粉和产卵,又浪费了聚果榕小蜂身上花粉和体内的虫卵,对榕树和小蜂繁殖后代均不利。
2.3 非传粉小蜂的瘿花对榕树种子形成的影响聚果榕果内小花不但为聚果榕小蜂提供繁衍生息的场所,也被其他非授粉小蜂所利用。所不同的是,聚果榕小蜂在果腔内产卵繁殖,其他非传粉小蜂则在榕果外面产卵。因此,非传粉小蜂一般具有较长的产卵器。并且,非传粉小蜂产卵活动多发生在聚果榕小蜂产卵之后。在聚果榕果内繁殖的非传粉小蜂经试验和观察发现有2个类群:一是造瘿类群,该类群属于膜翅目长尾小蜂科(Torymidae),种类为Platyneura testacea, Platyneura mayri,Platyneura agraensis, Platyneura sp.,共4种。它们主要寄生已经被聚果榕小蜂传粉受精的长柱花子房或已被聚果榕小蜂产卵寄生的短柱花子房,是严重影响聚果榕种子形成的一类害虫(图 2)。从图 2可以明显地看出,每一个榕果内只要造瘿类群的小蜂数量增多,聚果榕的种子数量就明显下降;当该类蜂数量降低时,种子数量又上升。

|
图 2 非传粉小蜂造瘿类群数量对聚果榕种子数量的影响 Fig. 2 Effect of gall maker number of nonpollinating fig wasps on seed number of F.racemosa |
另一个类群为复寄生(重寄生)类群,在聚果榕内仅发现属膜翅目刻腹小蜂科(Ormyridae)的1个种Apocrypta westwoodi,专门寄生在聚果榕小蜂幼虫或以上4种造瘿类的非传粉小蜂幼虫体内,靠寄生取食这些幼虫体内物质发育成长。当这种小蜂以造瘿类小蜂幼虫为主要寄生对象时,将减少造瘿类小蜂种群,对聚果榕小蜂种群和榕树种子形成有利;但当它以聚果榕小蜂为主要寄生对象时,对聚果榕种子形成造成间接的不利影响,因为聚果榕小蜂被寄生后种群数量下降,会对聚果榕小蜂后来的传粉造成严重影响。
3 讨论研究结果证明,聚果榕小蜂进聚果榕果内,结果表现为2方面:第一,对长柱小花传粉受精后产生榕树种子,使聚果榕能正常进行有性繁殖;第二,在短柱花内产卵形成瘿花,产生榕小蜂后代,植食性,最后发育为聚果榕小蜂,并作为榕树传粉的媒体。不同季节内,各种小花的比率不同。旱季的1月和11月,聚果榕产生的种子量最多,雨季末期的9—10月份次之,产生种子量最低的是3月份。旱季和雨季,瘿花的寄生比率没有显著差异,但不同月份之间差别较大,寄生率最高在旱季的1月份和雨季初期5月份,最低是在3月份。长花柱雌花和短花柱雌花的败育率,在气温最低的月份(1—2月)和雨量最多的月份(7—8月)结的榕果内最高,在雨季初期(5月)和旱季初期(11月)结的榕果内最低。聚果榕果内小花败育主要是受低温、多雨的气候影响。除聚果榕小蜂外,榕果内还有造瘿类和重寄生2类非传粉小蜂,它们对聚果榕的种子形成具有一定的影响。
聚果榕果内瘿花的产生直接和小蜂的产卵行为有关,小蜂(聚果榕小蜂和其他非传粉小蜂)的产卵行为是一个比较复杂的问题。为什么寄生在聚果榕果内的几种非传粉小蜂不象聚果榕小蜂一样,钻进榕果腔内产卵繁殖,而是用长的产卵管从果皮外表插入产卵?这些小蜂的产卵管是如何分辨形成种子的子房和被聚果榕小蜂寄生的瘿花子房的?这些小蜂的繁殖行为特性和机理有待于更多的研究、试验和观察。
聚果榕种子的产生不但受气候的影响,还与聚果榕传粉小蜂和非传粉小蜂的活动程度有关,同时也受到榕树所能提供的能量的限制。榕果内种子和瘿花比率如何分配?每一种非传粉小蜂对榕树繁殖率和聚果榕小蜂群体的影响程度等还需深入研究加以验证。
马炜梁, 陈勇, 李宏庆. 1997. 榕树及其传粉者研究综述. 生态学报, 17(2): 09-215. |
彭艳琼, 杨大荣, 周芳, 等. 2003. 木瓜榕传粉生物学. 植物生态学报, 27(1): 111-117. DOI:10.3321/j.issn:1005-264X.2003.01.017 |
王秋艳, 杨大荣, 彭艳琼. 2003. 西双版纳鸡嗉果榕小蜂繁殖和传粉行为. 昆虫学报, 46(1): 7-39. DOI:10.3969/j.issn.1005-0507.2003.01.002 |
吴征镒. 1995. 云南植物志(第6卷). 北京: 科学出版社, 595-671.
|
许再富. 1994. 榕树——滇南热带雨林生态系统中一类关键植物. 生物多样性, 2(1): 21-23. DOI:10.3321/j.issn:1005-0094.1994.01.004 |
徐磊, 杨大荣, 彭艳琼, 等. 2003. 西双版纳聚果榕榕果内小蜂群落结构及种间关系. 生态学报, 23(8): 1554-1560. DOI:10.3321/j.issn:1000-0933.2003.08.013 |
杨大荣, 王瑞武, 宋启示, 等. 2000. 西双版纳热带雨林聚果榕小蜂季节性变化规律. 林业科学研究, 13(5): 477-484. DOI:10.3321/j.issn:1001-1498.2000.05.004 |
杨大荣, 赵庭周, 王瑞武, 等. 2001. 西双版纳热带雨林聚果榕小蜂传粉生态学研究. 动物学研究, 22(2): 125-130. DOI:10.3321/j.issn:0254-5853.2001.02.008 |
杨大荣, 彭艳琼, 宋启示, 等. 2002a. 西双版纳热带雨林对叶榕传粉生物学. 植物学报, 44(5): 519-526. |
杨大荣, 彭艳琼, 张光明, 等. 2002b. 西双版纳热带雨林榕树种群变化与环境的关系. 环境科学, 23(5): 9-35. |
杨大荣, 彭艳琼, 张光明, 等. 2003. 西双版纳热带雨林聚果榕榕果的昆虫群落结构与多样性. 应用生态学报, 14(10): 1710-1714. DOI:10.3321/j.issn:1001-9332.2003.10.026 |
Berg C C. 1989. Classification and distribution of Ficus. Experientia, 45: 605-611. DOI:10.1007/BF01975677 |
Galil J, Eisikowitch D. 1968. On the pollination ecology of Ficus sycomorus in east Africa. Ecology, 49(2): 259-269. DOI:10.2307/1934454 |
Herre E A. 1989. Coevolution of reproduction characteristics in 12 species of New World figs and their pollinator wasps. Experientia, 45: 637-647. DOI:10.1007/BF01975680 |
Janzen D H. 1979. How to be a fig. Ann Rev Ecol Syst, 10: 13-51. DOI:10.1146/annurev.es.10.110179.000305 |
Ramirez W B. 1970. Host specificity of fig wasps (Agaonidae). Evolution, 24: 680-691. DOI:10.1111/j.1558-5646.1970.tb01804.x |
West S A, Herre E A. 1994. The ecology of the New World fig parasitizing wasps Idarnes and implications of the evolution of the fig-pollinator mutualism. Proc R Soc London Ser B, 258: 67-72. DOI:10.1098/rspb.1994.0143 |
Weiblen G D. 2002. How to be a fig wasp. Annual Review of Entomology, 47: 299-330. DOI:10.1146/annurev.ento.47.091201.145213 |
Wiebes J T. 1979. Coevolution of figs and their pollinators. Ann Rev Ecol Syst, 10: 1-12. DOI:10.1146/annurev.es.10.110179.000245 |