 2004, Vol. 40
2004, Vol. 40文章信息
- 王华田, 邢黎峰, 马履一, 孙鹏森.
- Wang Huatian, Xing Lifeng, Ma Lüyi, Sun Pengsen.
- 栓皮栎水源林林木耗水尺度扩展方法研究
- Study on Scaling-Up Method for Stand Water Consumption of Quercus variabilis Water Conservation Forest
- 林业科学, 2004, 40(6): 170-175.
- Scientia Silvae Sinicae, 2004, 40(6): 170-175.
-
文章历史
- 收稿日期:2003-08-08
-
作者相关文章



2. 北京林业大学资源与环境学院 北京 100083;
3. 中国林业科学研究院森林生态环境与保护研究所 北京 100091
2. College of Resources and Environment, Beijng Forestry University Beijing 100083;
3. Research Institute of Forest Ecology, Environment and Protection, CAF Beijing 100091
北京是一个严重缺水的城市,年均可供利用的水资源总量为400亿~420亿m3,人均350 m3,为世界平均水平的1/3,为全国的1/7。水资源短缺已经成为制约北京城市发展和社会事业进步的主要因子。北京市辖区内现有各类林业用地约90万hm2,其中以水源保护林为主的有林地46万hm2,主要树种为栎类(Qurcus)、油松(Pinus tabulaeformis)、刺槐(Robinia pseudoacacia)、山杨(Populus davidiana)、侧柏(Platycladus orientalis)等,对涵养和保护北京市水资源发挥着巨大作用。袁嘉祖(1999)利用线性规划方法,按照国民经济发展对水资源合理分配提出的要求和北京市水源保护林发展战略,根据现有森林耗水强度估算,提出北京市水资源能够承载的森林覆被率为24.16%(40万3 600 hm2),这显然不能满足北京市生态城市建设和绿色奥运的要求。如何准确计算和评价林木个体与群体的耗水量,进而选择低耗水树种和科学配置林分结构,对于编制水源保护林发展规划和营建技术方案无疑是十分重要和亟待解决的问题。
传统的林木个体与群体耗水计算方法普遍存在着精度偏低和描述能力不足的问题(王华田,2002)。Marshall (1958)、Swanson(1994)和Edwards等(1996)在Huber(1935)工作的基础上,成功地研制出第一代完整的树木边材液流检测系统。Granier (1987)根据双热电偶检测热耗散原理开发出热扩散式液流探针(thermal dissipation sap flow velocity probe,TDP),实现了连续或任意时间间隔树木边材液流速率的测定,能够动态地掌握整株树木的蒸腾耗水规律,并揭示其生态、生理作用机制(Wullschleger et al., 1998)。依据热平衡原理测定树木整株耗水特性,其变量只有检测部位的树干边材横断面积(Granier, 1987)。据此,可以根据林分群体边材分布规律,完成林分群体蒸腾耗水的尺度转换。Ladefoged(1963)、Cermak等(1987)、Werk(1988)、Hatton等(1990)、Vertessy等(1997)分别以冠幅、树干基部横断面积和林冠投影面积为基准变量,进行单木耗水的尺度扩展研究,结果都不甚理想。Vertessy等(1995)和Hatton等(1995)研究发现,叶面积是实现单木到林分耗水尺度转换最为可信的变量,认为木质部输导组织断面积、叶面积、胸径和基于生态地域理论的单木占地面积都是比较理想的空间推导纯量,其中树干胸径因为宜于测定,并且与比叶面积、边材面积等相关性强,是实现林木耗水尺度扩展的理想变量(孙鹏森等,2002)。
邢黎峰等(1998)通过对树木多种生长过程的研究,认为用Richards生长方程描述胸径、树高、材积等方面的生长特征,具有可塑性强、弹性大、描述准确、方程生物学意义完整等优点。树木单株累计日耗水过程为一典型的S曲线1),符合生物生长过程的一般特征,利用生长方程描述这一过程,通常可以取得满意的结果。
1) 王华田.北京市水源保护林区主要树种耗水特性研究.北京林业大学博士学位论文,2002
1 试验地点概况试验地位于北京林业大学鹫峰教学实习林场(39°54′N,116°28′E)。气候属于暖温带半湿润半干旱季风气候,年均温7~10 ℃,年降水量638.8 mm,蒸发量1 800~2 000 mm。土壤有褐土型耕作土、淋溶褐土、棕壤等类型。森林类型均属于防护林和风景林。自然植被属于典型温带落叶林带的山地栎林和油松林带。人工植被低山主要为侧柏林、刺槐林,中山地带为油松林、栎林和五角枫(Acer mono)与栓皮栎(Qurcus variabilis)混交林,高山地带有小片落叶松(Larix principis-rupprechtii)林。
试验地设在燕子岭栓皮栎人工林内,海拔480 m,坡度18°,梯田整地,淋溶褐土,土层厚度60 cm,枯落层3~15 cm。栓皮栎48年生,平均胸径19.7 cm,平均树高13.5 m。
2 试验材料与方法在20 m×30 m标准地上进行每木检尺,测量胸径(D1.3)、树高、冠幅(EW×NS)、枝下高等,计算林分平均木和径阶平均木的胸径和树高,并在标准地内标出,用于测定径阶单木整株耗水量。用生长锥法调查林内立木不同树干方位边材分布规律,同时选择与标准木生长状况相近的立木,伐倒后按照标准尺寸截取圆盘,查数年轮,测定边材和心材年轮数和年轮宽度,计算边材面积和心材面积,用于对生长锥调查结果的校正。
利用TDP探针于9月份连续测定栓皮栎人工林各径阶标准木树干边材液流速率,根据边材面积计算单木边材液流通量。利用微型自动气象站组成的测定系统连续测定林分大气和土壤常规环境因子。边材液流速率、单木液流通量、单木整株耗水量、气象因子和土壤因子的测定和计算方法见王华田等(2002)。
3 结果与分析 3.1 林分单木直径与边材面积的相关性表 1是栓皮栎人工林调查结果,相关分析结果表明,各径阶测定样木胸径与边材面积相关显著(R=0.915),但胸径与边材宽度、树冠面积与边材宽度、树冠面积与边材面积之间相关性不显著。
|
|
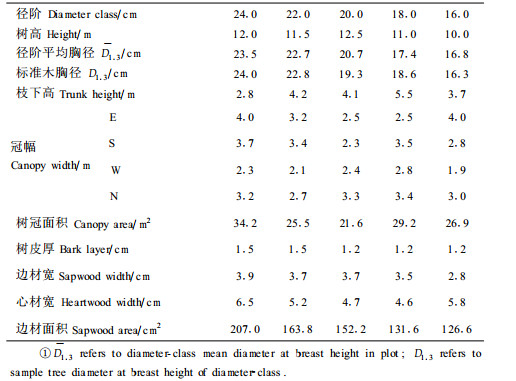
从图 1可以看出,栓皮栎边材液流速率和液流通量随着胸径的增大逐渐增大,不同直径日周期内树干边材液流进程表现出相似的规律,边材液流从8:00—9:00开始大幅度上升,11:00达到最大值,然后逐渐下降,18:00以后进入夜间液流缓慢传输阶段。

|
图 1 栓皮栎不同径阶树干边材液流速率和液流通量密度的日变化 Fig. 1 Diurnal flutuation of sapflow velocity and sapflow density in different diameter classes of Q.variabilis in Sep.15th |
图 2是不同直径栓皮栎单木日周期内各时段耗水量,从中可以看出,不同直径单木耗水的时间变化特征十分明显。边材液流的启动时间随直径增大而提前,上升速率随直径增大而增大。直径24.0 cm的单株,树干边材从凌晨4:00即有明显的液流传输,8:00突然增大,18:00以后液流传输进入低谷;直径22.8 cm的单株,边材液流从8:00开始上移,9:00开始大幅度上升,17:00停止;直径19.3 cm和18.6 cm的单株,液流开始传输和停止的时间相同,均为8:00和18:00,18:00以后进入缓慢的夜间液流传输阶段;直径16.3 cm单株液流开始传输的时间为9:00,在5个径阶中启动最晚,上升的幅度也最小。

|
图 2 栓皮栎不同径阶日耗水量变化进程 Fig. 2 Daily single-tree progressive water consumption in different diameter classes of Q.variabilis in Sep.15th |
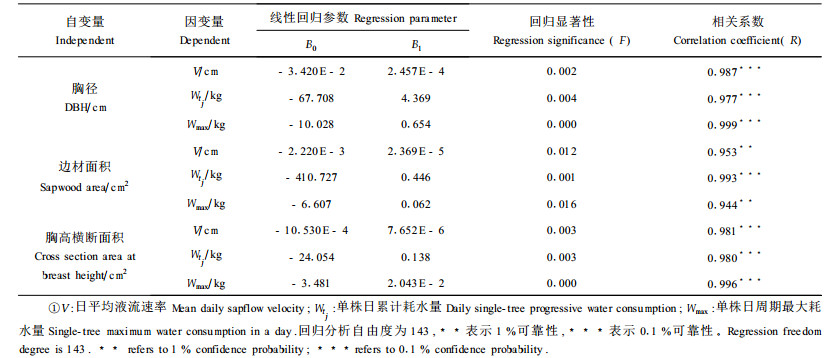
对栓皮栎胸径或树干横断面积与单株耗水量关系进行线性回归分析(表 2),结果发现,胸径、边材面积和胸高横断面积与单木日累计耗水量、日平均边材液流速率和日周期最大耗水量之间存在极显著的相关性,各变量之间的线性回归方程经过检验均达到极显著水平。其中特别有利用价值的是胸径和单木日耗水量的线性回归模型,根据这一简易线性模型,利用林分径阶分布规律可以直接在群体水平上对林分蒸腾耗水进行估测,实现由单木水平到群体水平耗水的尺度扩展。
|
|
树木边材液流研究的最终目的是研究树木的蒸腾耗水性,评价和比较不同树种、不同立地条件下、不同林分结构和林分不同生长发育阶段的蒸腾耗水量,并据此指导水源保护林造林树种选择和林分结构的合理配置。要实现这一目标必须解决耗水尺度的扩展问题。
图 3是根据各径阶日周期不同时段耗水量累加值绘制的栓皮栎日周期累计耗水动态曲线。从其变化趋势看,栓皮栎的累计耗水过程具有典型的生物生长曲线特征。据此确定选用Richar ds模型为基本模型,选用径阶和时间为自变量,对栓皮栎日周期耗水量作三维尺度展开,建立单木日周期耗水模型。Richards生长方程的一般表达形式为:

|
图 3 栓皮栎不同径阶单株累计耗水量日变化 Fig. 3 Daily sample tree progressive water consumption in diameter classes of Q.variabilis in Sep.15th |
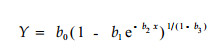
|
(1) |
式中:Y为时间t时刻生物累计生长量;b0是累积生长的饱和值,是生物学过程时间序列的最终稳定值,即生物学上限值;b1是生长初始值参数,决定着生长初始值的大小;b2是生长速率参数;b3是异速生长参数,它决定着S曲线的形状。
树木整株耗水量的时间累积过程具有生物学生长方程的一般特征,利用Richards生长模型描述林分内各径阶单木耗水量的时间分布过程可以得到理想结果。图 4是对图 3中栓皮栎人工林单木累计耗水量沿径阶变化的展开形式,相应的曲面特征方程为:

|
图 4 栓皮栎人工林Richards径阶耗水量模型的空间曲面特征 Fig. 4 3-D curve of Richards model of diameter class water consumption of Q.variabilis stand |
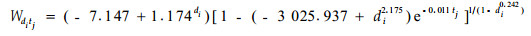
|
(2) |
式中:Wditj为林分中胸径di的栓皮栎单株在时间tj时刻的累积耗水量;参数b0=-7.147+1.174di,b1=-3 025.937+di2.175,b2=0.011,b3=di0.242。采用参数回收算法同步求解方程参数,相关系数R=0.985 8。
将径阶范围内任一单株的胸径和相应的时间(min)代入方程(2)中,即可得到该单株日周期内自早6:00至给定时间的累计耗水量。对该方程求导,并令二阶导数和三阶导数为0,可以得到2组特征值。第1组为曲线的拐点q,即单木日周期耗水速率最大值及其出现时刻;第2组是曲线上单木耗水速率由慢变快和由快变慢的分界点,它们位于曲线拐点的上、下方,其中位于拐点下方的是曲线上耗水速率由慢变快的凹点p1,位于拐点上方的是曲线上耗水速率由快变慢的凸点p2。
方程拐点坐标q(x, y)为:
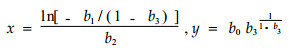
|
(3) |
耗水速率转换点p1(x1, y1)和p2(x2, y2)的坐标为:
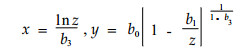
|
(4) |
其中
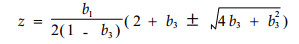
|
(5) |
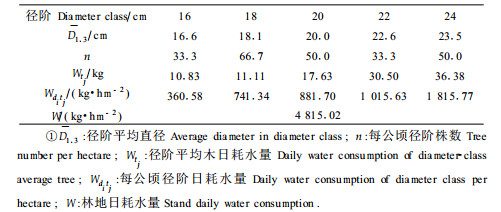
根据标准地调查结果,利用耗水方程计算被测林分栓皮栎的径阶耗水量分布,结果为:径阶16、18、20、22、24 cm(每公顷株数分别为33.3、66.7、50.0、33.3、50.0株),径阶日耗水量分别为360.58、741.34、881.70、1 015.63、1 815.77 kg,每公顷混交林中栓皮栎日耗水总量4 815.02 kg(表 3)。
|
|
对栓皮栎林分内不同径阶单株的树干边材液流速率同时进行测定,发现树干直径与边材面积、以及树干直径或冠幅与边材液流速率、单株耗水量之间存在着显著的相关性,与Vertessy(1995)、孙鹏森等(2002)、刘奉觉等(1997)的研究结果一致,符合孙鹏森等(2002)提出的实现林木耗水尺度扩展所需要的纯量要求,具有易于调查、直观、可操作性强等优点,因而选择胸高直径作为林木耗水尺度扩展的变量是可行的。
林木耗水尺度扩展过程中存在2个难点:第1个难点是如何实现由单木耗水量沿时间进程和径阶的扩展,其中变量的选择及其与单木耗水量的关系、模型的选择和方程的求解是尺度扩展的核心,其难度大于一般林木生长模型和人工林经营模型(杜纪山等, 1997)。单木日周期累计耗水量的研究发现,其日周期变化进程呈S形,具有典型的生物生长曲线特征(王华田等, 2002;孙鹏森等, 2002),利用Richards生长模型拟合效果理想(邢黎峰等, 1998)。树干直径、边材面积和单株耗水量的相关分析结果也证明树干直径与边材面积、树干直径与单株耗水量之间有非常好的相关性。为此确定采用树干胸径这一最常用的测树指标作为单木-群体耗水量尺度扩展变量。对栓皮栎不同径阶单株耗水量日变化的测定结果进行尺度扩展,所得到的单木三维耗水模型拟合效果十分理想。耗水尺度扩展中存在的第2个难点是在对时间尺度和径阶尺度三维扩展的基础上如何将主导环境因子加入到模型中,即如何利用现实林分环境条件下林分群体耗水量测定结果到任意环境条件下林分耗水量的尺度扩展问题。由于影响林分蒸腾耗水的环境因子种类多,作用机理复杂,且彼此之间交互作用变化不定,尺度扩展中面临的问题庞大而复杂,难以短时间得到解决。
通常情况下,山地水源保护林林分水量平衡方程中的收入项仅为年度降水量,而支出项主要为林冠蒸腾、林冠降水截流和林地地表蒸发。对于特定树种的林分而言,在一定气候条件下的林冠截流和地表蒸发与降水量相关密切(温远光等, 1996)。依据生长季节林地土壤水分状况适宜条件下山地半阴坡上位中层土48年生栓皮栎林分群体耗水三维模型推算,600 mm降水量林地水分环境容量的最大承载力(即林分理论生态密度)为1 243株·hm-2。此时林地的水分环境容量与林分蒸散耗水量相同,林地水分状况能够满足树木生长的需要,但没有多余径流产出;林分密度超过这一临界值,树木生长由于得不到充足的水分供应而受到一定程度的限制;林分密度小于这一临界值,则在林木正常生长的前提下,林地尚有多余径流产出(忽略土壤深层下渗),径流产出量随着林分密度的减小而增加。
杜纪山, 唐守正. 1997. 林分断面积生长模型研究评述. 林业科学研究, 10(6): 599-606. |
刘奉觉, 郑世锴, 巨关升. 1997. 树木蒸腾耗水测算技术的比较研究. 林业科学, 33(2): 117-126. DOI:10.3321/j.issn:1001-7488.1997.02.003 |
马履一, 王华田. 2002. 油松、侧柏深秋边材木质部液流变化规律的研究. 北京林业大学学报, 23(4): 23-37. DOI:10.3969/j.issn.1671-6116.2002.04.004 |
孙鹏森, 马履一. 2002. 水源保护树种耗水特性研究与应用. 北京: 中国科技文献出版社.
|
王华田, 马履一. 2002. 利用热扩散式边材液流探针(TDP)测定树木整株蒸腾耗水量的研究. 植物生态学报, 26(6): 661-667. DOI:10.3321/j.issn:1005-264X.2002.06.005 |
温远光, 刘世荣.中国主要森林生态类型降水截留规律的数量分析.见: 蒋有绪.中国森林生态系统结构与功能规律研究.北京: 中国林业出版社, 1996: 23-34
|
邢黎峰, 孙明高, 王元军. 1998. 生物生长的Richards模型. 生物数学学报, 28(4): 460-464. |
袁嘉祖. 21世纪首都林业发展战略的思考.见: 21世纪的首都绿化.北京: 中国林业出版社, 1999
|
Cermak J, Kucera J. 1987. Transpiration of mature stand of spruce (Picea aies (L.) Karst.) as estimated by tree trunk heat balance method. Forest Hydrology and Watershed Management, 167: 311-317. |
Edwards W R N, Becker P. 1996. A unified nomenclature for sap flow measurements. Tree Physiology, 17: 65-67. |
Granier A. 1987. Evaluation of transpiration in a Douglas fir stand by means of sap flow measurements. Tree Physiology, 3: 309-319. DOI:10.1093/treephys/3.4.309 |
Hatton T J, Wu H I. 1995. Scaling theory to extrapolate individual tree water use to stand water use. Hydrol Proc, 9: 527-540. DOI:10.1002/hyp.3360090505 |
Hatton T J, Vertessy R A A. 1990. Transpiration of plantation Pinus radiata estimated by the heat pulse method and the Bowen Ratio. Hydrological Processes, 4: 289-298. DOI:10.1002/hyp.3360040309 |
Huber B. 1935. Beobachtung und Messung pflanzlicher Safrströme. Berichte der Deutschen Botanischen Gesellschaft, 50: 89-109. |
Ladefoged K. 1963. Transpiration of forest trees in closed stand. Physiologia Plantarum, 16: 378-414. DOI:10.1111/j.1399-3054.1963.tb08319.x |
Marshall D C. 1958. Measurement of sap flow in conifers by heat transport. Plant Physiology, 33: 385-396. DOI:10.1104/pp.33.6.385 |
Swanson R H. 1994. Water transpired by trees is indicated by heat pulse velocity. Agricultural Meteorology, 72: 113-132. DOI:10.1016/0168-1923(94)90094-9 |
Vertessy R A, Benyon R, O'Sullivan S K, et al. 1995. Relationship between diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiology, 15: 559-568. DOI:10.1093/treephys/15.9.559 |
Vertessy R A, Hatton T J, Reece P, et al. 1997. Estimating stand water use of large mountain ash trees and validation of the sap flow measurement technique. Tree Physiology, 17: 747-756. DOI:10.1093/treephys/17.12.747 |
Werk K S. 1988. Performance of Picea abies (L.) Karst stands at different stages of decline Ⅲ: Canopy transpiration of green trees. Oecologia, 76: 519-524. DOI:10.1007/BF00397863 |
Wullschleger S D, Meinzer F C, Vertessy R A A. 1998. Review of whole-plant water use studies in trees. Tree Physiology, 18(8/9): 499-512. |