 2004, Vol. 40
2004, Vol. 40文章信息
- 谈家金, 冯志新.
- Tan Jiajin, Feng Zhixin.
- 松材线虫与其伴生细菌在寄主内的种群动态
- Population Dynamics of Pine Wood Nematode and its Accompanying Bacterium in the Host
- 林业科学, 2004, 40(6): 110-114.
- Scientia Silvae Sinicae, 2004, 40(6): 110-114.
-
文章历史
- 收稿日期:2003-01-24
-
作者相关文章


2. 华南农业大学植物线虫研究室 广州 510642
2. Lab of Plant Nematology, South China Agricultural University Guangzhou 510642
松材线虫病是松树的一种毁灭性病害,该病于1905年在日本九洲岛的长崎暴发,当时认为是几种昆虫所致,未受到重视(Mamiya,1983)。直至1969年,日本Tokushige(1968)和Kiyohara(1971)首先提出该病与1种伞滑刃线虫有关,并通过大量接种试验证明了该线虫的致病性,后经Mamiya和Kiyohara鉴定为松材线虫(Bursaphelenchus xylophilus)后(Mamiya et al., 1972),才引起广泛深入的研究。该病涉及到寄主、线虫、媒介昆虫、真菌、细菌以及环境因素等多个方面,至今其致病机制尚未弄清。Oku等(1980)从松材线虫虫体和感病松树体内分离到1种致病细菌,可使松苗萎蔫,首次提出该病与细菌有关;Kawazu等(1996)提出该病的真正病原是松材线虫携带的致病细菌;赵博光等(2000a;2000b)提出该病是松材线虫和其携带致病细菌的复合侵染病害,并通过抗生素控制致病细菌的方法控制松材线虫病。可见,通过研究细菌与该病的关系,对弄清其致病机理和解决防治难题具有重要意义。前人主要利用感病松树的愈伤组织及组培苗作接种材料研究了细菌与病害的关系,本文选用马尾松(Pinus massoniana)水培离体松枝作接种材料,研究了接种后松材线虫与其伴生细菌在松枝内的种群动态。
1 材料与方法 1.1 松材线虫和细菌的获得及预处理松材线虫于1998年6月分离自广东省东莞市樟木头镇一濒死马尾松, 室内盘多毛孢(Pestalotia sp.)上培养繁殖。接种前,松材线虫经5 mg·mL-1硫柳汞消毒1 h,无菌去离子水洗3次。松材线虫伴生细菌坚强芽孢杆菌(Bacillus firmus)的获得及预处理见谈家金等(2001a)中的菌株GD1方法1)。
1) 谈家金.坚强芽孢杆菌与松材线虫病关系的研究.华南农业大学博士论文, 2001
1.2 松枝采集、水培和接种2001年4月从广州市林校的健康马尾松(胸径10 cm左右)上,采集1~2年生松枝,带回温室用自来水培养。松枝水培参照刘军民等(1995)的方法。采用截枝套管法(王玉嬿等,1985)接种。设3个处理,1个对照,每处理接种18个松枝。1)线虫+细菌:接种5 000条松材线虫和3×108CFU,坚强芽孢杆菌的混合液;2)线虫:接种5 000条松材线虫;3)细菌:接种3×108CFU坚强芽孢杆菌;4)对照:接无菌去离子水。
1.3 病情观察及记载 1.3.1 症状观察接种后每3天观察松枝的外部症状。各松枝的病情分级和病情指数的计算基本按谈家金等(2000)的方法进行。病情分0级:松枝正常,叶绿色;1级:1/4以下叶发黄;2级:1/4~3/4叶发黄;3级:3/4以上叶发黄,1/2以下叶枯萎;4级:1/2以上叶枯萎,松枝濒死或死亡。
1.3.2 松枝髓部观察接种后3、6、9、12、15 d,各处理取3个松枝,去除针叶并用刀片将松枝切成薄片后,观察接种枝、主枝下部和主枝中部的髓部颜色变化。
1.3.3 松材线虫种群动态接种后3、6、9、12、15 d各处理取3棵松枝,去除针叶后于松枝的各部分取5 cm小段,切成薄片后,分离松材线虫并计数。然后将分离后的材料烘干称重,算得各样的松材线虫数量(条·g-1)。各取样部位如下。1)接种枝:离主枝5~10 cm,即接种枝顶端5~10 cm一段;2)主枝下部:接种枝以下5~10 cm一段;3)主枝中部:接种枝以上5~10 cm一段;4)下部侧枝:离主枝5~10 cm一段;5)新梢:新梢中部5~10 cm一段。
1.3.4 细菌的检测接种后3、6、12、15 d,各处理取3棵松枝,取主枝中部0.5 cm小段,洗净,体积分数75%酒精浸1~2 s,火焰上烧去酒精后,无菌水浸洗3次。用镊子和手术刀将枝茎切成厚约1 mm,长宽约3~6 mm的薄片,放入营养肉汤培养基[NB,成分参照Kawazu等(1996)]中,黑暗下、27 ℃振荡培养后营养琼脂培养基[NA,成分参照Kawazu等(1996)]上划线分离。
2 结果 2.1 症状观察松材线虫和坚强芽孢杆菌混合接种,松枝发病,单独接种松材线虫或坚强芽孢杆菌,松枝不发病。接种后15 d,线虫+细菌处理的病情指数达0.50。此时,线虫和细菌处理的松枝正常(图 1)。

|
图 1 松枝的病情指数 Fig. 1 The disease index of the treated aquacultural excised branches of P. massoniana —◇—线虫+细菌Nematode+bacterium;—○—线虫Nematode;—△—细菌Bacterium;—×—对照CK.下同The same below. |
松材线虫和坚强芽孢杆菌混合接种,松枝髓部发生褐变,髓部褐变的过程是由接种枝发展到主枝,再由主枝下部向上部发展。接种6 d后,线虫+细菌处理的接种枝、主枝下部和主枝中部的髓部发生褐变。线虫、细菌2个处理和对照的松枝髓部正常(表 1)。
|
|
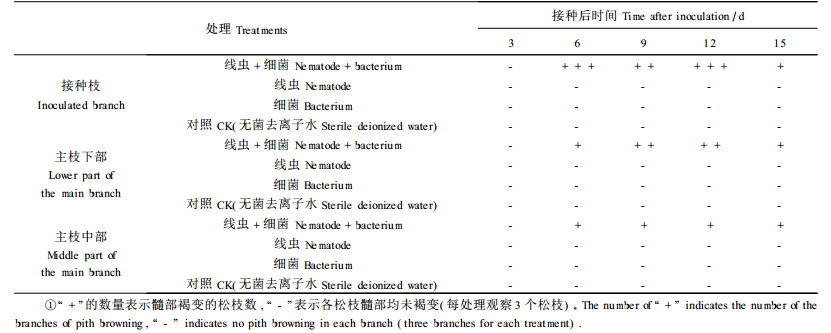
松材线虫和坚强芽孢杆菌混合接种较单独接种松材线虫,松枝中的松材线虫繁殖量大、扩散速度快;松材线虫在松枝中扩散的过程是:接种枝→主枝→侧枝→新梢,在主枝中由下部向上部扩散。接种枝中的松材线虫数量于接种后开始下降,随后上升,至一定数量后,又开始下降。线虫和坚强芽孢杆菌混合接种后3 d,接种枝中的松材线虫数量降为1 656条·g-1,随后一直上升,接种后12 d,松材线虫数量达3 232条·g-1,随后下降至1 591条·g-1;单独接种松材线虫后6 d,接种枝中的松材线虫数量降为352条·g-1,此后保持上升,接种后12 d,松材线虫数量达1 371条·g-1,随后下降至543条·g-1(图 2)。主枝下部于接种后3 d即可检测到松材线虫,此后数量增加,接种后12和15 d松材线虫数量增加明显。松材线虫和坚强芽孢杆菌混合接种较单独接种松材线虫,在主枝下部的松材线虫数量大。接种后15 d,前者的松材线虫数量为2 521条·g-1,而后者的松材线虫数量仅为463条·g-1(图 3)。主枝中部于接种后6 d开始检测到松材线虫,此后,松材线虫数量增加不明显,并出现波动。松材线虫和坚强芽孢杆菌混合接种与单独接种松材线虫,在主枝中部的松材线虫数量差异不明显。下部侧枝也于接种后6 d检测到松材线虫,但数量很少。松材线虫和坚强芽孢杆菌混合接种与单独接种松材线虫后,下部侧枝的松材线虫数量最高分别为3条·g-1和2条·g-1(图 4)。新梢于松材线虫和坚强芽孢杆菌混合接种后9 d,开始检测到松材线虫,随后松材线虫数量缓慢增加。至接种后15 d,松材线虫数量达8条·g-1。而单独接种松材线虫后,新梢一直未检测到松材线虫(图 5)。单独接种坚强芽孢杆菌和对照松枝中均未分离到松材线虫。

|
图 2 松枝接种枝的松材线虫数量变化 Fig. 2 Change of the number of B. xylophilus in the inoculated branch |

|
图 3 松枝主枝下部的松材线虫数量变化 Fig. 3 Change of the number of B.xylophilus in the tower part of the main branch |

|
图 4 松枝下部侧枝的松材线虫数量变化 Fig. 4 Change of the number of B.xylophilus in the lower lateral branch |

|
图 5 松枝新梢的松材线虫数量变化 Fig. 5 Change of the number of B.xylophilus in the current shoot of the branch |
松材线虫和坚强芽孢杆菌混合接种初期,细菌的检测量较大,而后期各处理的细菌检测量差异不明显。至接种后6 d,线虫+细菌处理的6个松枝中有5个检测到细菌,而线虫、细菌2个处理和对照分别只有1、0和2个松枝检测到细菌(表 2)。检测到的细菌,经鉴定均为坚强芽孢杆菌。
|
|

松材线虫和坚强芽孢杆菌混合接种,松枝发病,单独接种松材线虫或坚强芽孢杆菌,松枝不发病。从而笔者认为细菌和松材线虫均是松材线虫病不可缺少的致病因素,这与Kawazu等(1996;1997;1998)的观点有相似之处。本文的接种材料为马尾松水培离体松枝,试验结果能否适用于松苗或大树,尚有待进一步试验验证。文中消毒过的松材线虫经检验后仍带菌,这可能说明松材线虫侵入后,树体内致病细菌的数量直接决定着病害的发生与否。
松材线虫和坚强芽孢杆菌混合接种后,松枝髓部发生褐变,髓部褐变的过程是由接种枝发展到主枝,再由主枝下部向上部发展,而单独接种松材线虫或坚强芽孢杆菌与对照一样,松枝髓部未见褐变。本次试验进一步证实髓部褐变是松材线虫病的特征之一,并揭示了松材线虫病引起的寄主髓部褐变过程。
松材线虫和坚强芽孢杆菌混合接种较单独接种松材线虫,松枝中的松材线虫繁殖量大、扩散速度快。这是因为松材线虫和坚强芽孢杆菌共同侵入寄主后,较快地与寄主建立寄生关系,使得松材线虫迅速繁殖和扩散。松枝的外部症状出现前,松材线虫已扩散于整个松枝,但接种枝以外的松枝松材线虫数量很少。松材线虫在松枝中扩散的过程是:接种枝→主枝→侧枝→新梢,在主枝中由下部向上部扩散。由此可见,松枝髓部褐变过程和松材线虫在松枝中扩散过程相一致,但松材线虫扩散分布先行于髓部褐变。刘军民等(1995)研究表明,马尾松水培离体松枝接种松材线虫后,线虫向主枝基部扩散的数量和速度比向顶部的大,本次试验结果基本相同,但初步明确了松材线虫在寄主中的扩散过程。试验发现,离体松枝主枝含水量由下向上逐渐升高,至水培后期则逐渐降低,松材线虫在松枝中的扩散过程可能受松枝含水量的影响。
松材线虫和坚强芽孢杆菌混合接种初期,松枝中细菌的检测量较大,此时,单独接种松材线虫或坚强芽孢杆菌和对照松枝中很少检测到细菌。本次试验证实了松材线虫侵染早期,致病细菌在松树体内已经开始繁殖。Kuroda等(1992)指出,接种后第5周,细菌在接种树茎部才检测到。小林享夫等(1983)研究表明,自然感病松树松脂渗出异常时,木材中细菌的检出与健康松树的一致,检出率极低。这可能因为各研究者采用的细菌分离或检测方法不同,导致研究结果各异。今后应采取灵敏的细菌检测手段,深入研究松材线虫侵染后,松树体内细菌的变化。
松材线虫病早期诊断一直是该病的一个研究热点。从目前的研究情况看,流胶法虽对该病的早期诊断有一定的可靠性,但松树流胶停止后往往很难挽救,苯甲酸法和同工酶法应用于早期诊断还需大量试验验证(Kawazu,1990;刘伟等,1998;谈家金等,2001b)。本次试验发现在外部症状出现前,松枝的主枝下部和主枝中部髓部发生褐变,这表明通过寄主主干下部的髓部观察可对松材线虫病作出早期诊断。在本次试验中,松材线虫和坚强芽孢杆菌混合接种初期,松枝中细菌的检测量较大,这是否暗示可通过致病细菌数量的检测对病害进行早期诊断,尚需进一步研究。
刘军民, 冯志新. 1995. 松材线虫病组织病理学研究. 植物病理学报, 25(2): 171-174. DOI:10.3321/j.issn:0412-0914.1995.02.003 |
刘伟, 杨宝君, 徐福元, 等. 1998. 松材线虫病树早期诊断的研究Ⅰ.马尾松、黑松松材线虫病树的早期诊断. 林业科学研究, 11(5): 455-460. DOI:10.3321/j.issn:1001-1498.1998.05.001 |
谈家金, 王新荣, 冯志新. 2001a. 松材线虫伴生细菌与松树萎蔫病关系的初步研究. 植物检疫, 15(6): 326-328. |
谈家金, 吴慧平, 杨荣铮, 等. 2001b. 松材线虫对黑松几个生理生化指标的影响. 中国森林病虫, 20(4): 3-5. |
谈家金, 杨荣铮, 吴慧平. 2000. 不同地理种群的松材线虫对马尾松的致病力差异. 植物检疫, 14(6): 324-325. DOI:10.3969/j.issn.1005-2755.2000.06.002 |
王玉嬿, 石运琳, 周新胜, 等. 1985. 南京地区黑松萎蔫线虫病发病规律的初步观察. 森林病虫通讯, (2): 15-17. |
小林享夫, 佐佐木克彦. 1983. 与松材线虫病相关的根系中真菌和木材中真菌Ⅱ. 森林防疫, 32(10): 10-15. |
赵博光, 高蓉, 巨云为, 等. 2000a. 抗生素对松材线虫病的影响. 南京林业大学学报, 24(4): 75-77. |
赵博光, 郭道森, 高蓉, 等. 2000b. 细菌分离物B619与松材线虫病关系的初步研究. 南京林业大学学报, 24(4): 72-74. |
Kawazu K. 1990. Changes in constituents of pine wood by infection of pine wood nematode. Nippon Nogeikagaku Kaishi, 64: 1262-1264. DOI:10.1271/nogeikagaku1924.64.1262 |
Kawazu K, Kaneko N. 1997. Asepsis of the pine wood nematode isolate OKD-3 causes it to lost its pathogenicity. Japanese Journal of Nematology, 27(2): 76-80. DOI:10.3725/jjn1993.27.2_76 |
Kawazu K, Yamashita H, Kobayashi A, et al. 1998. Isolation of pine-wilting bacteria accompanying pine wood nematode, Bursaphelenchus xylophilus, and their toxic metabolites. Scientific Reports of the Faculty of Agriculture, Okayama University, 87: 1-7. |
Kawazu K, Zhang H, Yamashita H, et al. 1996. Relationship between the pathogenicity of the pine wood nematode, Bursaphelenchus xylophilus, and phenylacetic acid production. Biosci Biotech Biochem, 60(9): 1413-1415. DOI:10.1271/bbb.60.1413 |
Kiyohara T, Tokushege Y. 1971. Inoculation experiments of a nematode, Bursaphelenchus sp. onto pine trees. J Jpn For Soc, 53: 210-218. |
Kuroda K, Ito S. 1992. Migration speed of pine wood nematode and activities of other microbes during the development of pine-wilt disease in Pinus thunbergii. J Jpn For Soc, 74(5): 383-389. |
Mamiya Y. 1983. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Ann Rev Phytopathol, 21: 201-220. DOI:10.1146/annurev.py.21.090183.001221 |
Mamiya Y, Kiyohara T. 1972. Description of Bursaphelenchus lignicolus n. sp. (Nematoda: Aphelenchoididae) from pine wood and histopathology of nematode-infested trees. Nematologica, 18: 120-124. DOI:10.1163/187529272X00296 |
Oku H, Shiraishi T, Ouchi S, et al. 1980. Pine wlit toxin, the metabolite of a bacterium associated with a nematode. Naturwissenschaften, 67: 198-199. |
Tokushige Y, Kiyohara T. 1969. Bursaphelenchus sp. in the wood of dead pine trees. J Jpn For Soc, 51: 193-195. |