 2004, Vol. 40
2004, Vol. 40文章信息
- 翟明普, 蒋三乃, 贾黎明.
- Zhai Mingpu, Jiang Sannai, Jia Liming.
- 杨树刺槐混交林细根养分动态研究
- Study on Fine-Root Nutrient Dynamics in the Mixed Plantation of Poplar and Black Locust
- 林业科学, 2004, 40(4): 46-51.
- Scientia Silvae Sinicae, 2004, 40(4): 46-51.
-
文章历史
- 收稿日期:2002-10-11
-
作者相关文章


树木细根在森林生态系统的养分循环和能量流动中起着重要作用,虽然只占总生物量的5%左右,但其生长量可占森林总初级生产力的50%~75%(Hendrick et al., 1992;Nadelhoffer et al., 1992)。细根死亡是有机质和养分元素向森林土壤归还的重要途径,通过根系死亡归还到土壤中的N素比地上凋落物多18%~58%(McClaugherty et al., 1982;Vogt et al., 1986)。随着人们对细根在森林生态系统养分循环中作用的重视,近20多年来,细根研究已成为森林生态系统研究中的一个热点,在国外受到广泛的关注(Nadelhoffer et al., 1992;Finer et al., 1997;Thorup-Kristiina et al., 1998;Robinson et al., 1999;Roy & Singh,1995;Ruess et al., 1996),但目前国内关于树木细根的研究报道不多(单建平等,1993;廖利平等,1999;2000;翟明普等,2002)。由于细根的周转与分解在养分循环方面具有重要的作用,而混交林中不同树种的细根周转及其养分归还又有各自不同的规律,这必然会影响其种间相互关系。对不同树种纯林与混交林细根动态及其养分状况的深入研究能更清楚地揭示混交林的种间养分关系。
杨树(Populus)-刺槐(Robinia pseudoacacia)混交林是一种典型的固氮树种和非固氮树种组成的混交林,在我国北方沙地造林中具有重要意义。前期研究表明,其生产力高和生态协调能力强的主要原因是两树种的养分互补(沈国舫等,1998;贾黎明等,1997;翟明普等,1997)。本文试图通过细根分解和养分变化等方面的研究,更深入地揭示杨树刺槐混交林的养分协调机制。
1 实验地概况实验地位于北京市共青林场郝家疃分场,位于潮白河畔,116°46′E,40°16′N,海拔30 m。共青林场于1962年建场,面积为980 hm2。土壤主要为冲积沙土,少量含有壤质夹层,地下水位5 m左右;有机质含量低于5.0 g·kg-1,蓄水保肥能力差。本地区属暖温带大陆性气候,春季干旱多风,夏季炎热多雨,年均气温11.4℃,积温5 400℃。年均降水量630 mm,主要集中于7、8月份,年均蒸发量为2 000 mm;无霜期180—200 d。
2 研究方法 2.1 研究对象研究对象为北京潮白河林场的加杨(P. canadansis)纯林、刺槐纯林、加杨和刺槐的混交林。加杨27 a生,刺槐24 a生,纯林株行距为3 m×3 m,杨树刺槐行状混交,行距3 m,杨树刺槐行距分别为1.5 m,调查时混交林中刺槐保存率52.6%。
2.2 实验布置及测定方法在不同林分各选面积为0.2 hm2的典型样地。于2000年2月用内径为6 cm的土钻在样地内取样,拣根,带回室内在1 mm网筛上冲洗,按树种和径级分拣,2 mm者为细根。将细根风干后剪成3 cm的根段,留下部分根样作含水率和初始养分分析用,其余作为试验样品。试验根样按树种装入网眼为0.5 mm的尼龙网袋内。杨树细根、刺槐细根单独分解试验每袋5 g,杨、槐细根按1:1的重量比混合。装好袋后,将杨树和刺槐细根分别放入纯林样地,混合细根放入混交林样地。分解袋放入离地表 10 cm的土壤中,每隔1个月取样1次,每次5袋,取回后立即清除泥土等非根系物质,80℃烘干至恒重,粉碎作养分含量分析。元素含量测定方法:N素用凯氏定氮法,P素用钼锑抗比色法,K、Ca、Mg用火焰光度法。
2.3 养分归还量计算养分归还量=(细根年死亡量×最初养分含量)-(分解残留量×残留物养分含量)。
3 结果与讨论 3.1 细根分解与养分释放细根分解过程中常常伴随着物质淋溶、迁移,土壤微生物代谢对养分元素的吸收与释放等过程,与外界环境进行物质交换。因此,细根分解过程中元素含量也在不断发生变化,有些元素以积累为主,有些则以释放为主,或积累与释放过程交替进行。同时,细根在分解过程中养分含量的变化也与供试根系本身的物理结构、化学组成相关。
从细根的初始养分元素浓度(表 1)看出,杨树细根中的养分元素P和Mg的浓度比刺槐细根高,而N、K、Ca的浓度都较刺槐细根低;杨、槐混合细根的各养分元素浓度介于两者之间。与两树种地上枯落物相比,杨、槐细根的N、P、Ca、Mg浓度都较前者高,尤其是N素,杨树细根、槐细根以及杨槐混合细根分别为杨树、及两者混合枯落物的1.74、1.84和1.89倍,只有K的浓度较前者低,这一结果与前人的研究结论类似(Vogt et al., 1986)。
|
|
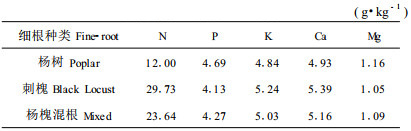
结果表明,细根分解过程中N、P、K、Ca、Mg元素含量是变化的(图 1~5)。图 1说明,杨树细根在自身纯林和刺槐纯林下300 d分解过程中,一直处于N素积累过程,这与其N素含量低而C/N比高的特性有关;刺槐细根在杨、槐纯林下分解初期的N素浓度比较稳定,但在自身纯林下分解60 d后N素浓度开始下降,而在杨树纯林下分解120 d后开始下降,可见刺槐细根在分解试验期内处于N素释放过程,而且在刺槐纯林下分解时的释放速率比在杨树纯林下速度快;杨、槐混合细根在混交林下分解中,N素浓度持续下降,120 d后降低的速度加快。可见与刺槐细根混合,可加速杨树细根中N素的释放过程,有利于林地死细根N。图 2说明,杨树细根在自身纯林和刺槐纯林下分解中,P素浓度持续下降,但在杨树纯林下分解降低的幅度小。刺槐细根在自身纯林和杨树纯林下分解过程中P素浓度变化规律与杨树细根类似,也处于养分释放状态。180 d以前刺槐细根在两种林分下P浓度均呈降低趋势,但180 d以后,刺槐纯林下分解的刺槐细根中P素浓度降低速度加快。杨槐细根混合分解P素也处于释放状态,其释放速率较杨树细根单独分解慢而比刺槐细根单独分解快。

|
图 1 杨、槐细根分解过程中N素浓度的变化 Fig. 1 Change of N concentration in decomposing fine root —◃—杨根—杨树纯林Poplar root in poplar stands; —□—杨根—刺槐纯林Poplar root in Black Locust stands; —△—槐根—杨树纯林Black Locust root in poplar stands; —×—槐根—刺槐纯林Black Locust root in Black Locust stands; —○—杨槐混根—混交林Black Locust and poplar root in mixed stands。下同。The same below. |

|
图 2 杨、槐细根分解过程中P素浓度的变化 Fig. 2 Change of P concentration in decomposing fine root |

|
图 3 杨、槐细根分解过程中K素浓度的变化 Fig. 3 Change of K concentration in decomposing fine root |

|
图 4 杨、槐细根分解过程中Ca素浓度的变化 Fig. 4 Change of Ca concentration in decomposing fine root |

|
图 5 杨、槐细根分解过程中Mg素浓度的变化 Fig. 5 Change of Mg concentration in decomposing fine root |
细根分解过程中K素养分浓度变化如图 3所示。杨树细根在自身纯林和刺槐纯林下分解时K素浓度变化趋势一致,都处于释放状态,但在分解初期浓度降低很快,到180 d时几乎下降了80%,后120 d释放速度放慢;刺槐细根在纯林下分解时,K素浓度变化规律与杨树细根类似,但释放速度较杨树细根稍慢,而且在刺槐纯林下分解时浓度下降幅度较在杨树纯林下稍慢;杨、槐细根混合分解,K素释放速率介于杨、槐细根单独分解之间。
图 4说明,杨树细根在自身纯林和刺槐纯林下分解过程的初期(60 d以前),Ca元素浓度小幅度下降,随后上升,而且一直延续到试验结束时,最初的养分浓度下降可能是淋溶作用的结果;刺槐细根在分解过程中Ca素浓度一直在增大,而且在其自身纯林下分解时升高的速率比在杨树纯林下稍快;杨、槐细根在混交林下混合分解时Ca素也呈升高趋势,但速度缓慢。
图 5看出杨、槐细单独分解和混合分解过程中,Mg元素的浓度持续升高,杨、槐细根单独分解时升高的速率基本相同,只是杨树细根在自身纯林下分解180 d后升高的速度有所加快,而刺槐细根在两种纯林下以及杨树细根在刺槐纯林下分解时升高的趋势比较平缓;杨、槐细根混合分解时Mg元素浓度一直在增大,增高的速度介于杨、槐细根单独分解之间。
3.1.2 细根分解过程中的元素重量损失试验结果(表 2)表明,在分解至300 d时,细根单独分解和混合分解条件下都表现为养分释放(重量损失百分率大于0),但不同条件下释放程度不同。细根分解过程中N素的释放规律差异较大:杨树细根N素的重量损失百分率最低,只有23%~24%,而且在自身纯林下和在刺槐纯林下单独分解的重量损失率几乎相等;刺槐细根的N素损失百分率比杨树细根高得多,达68%~75%,且在自身纯林下分解的重量损失率比在杨树纯下分解有所提高;杨、槐细根在混交林下混合分解的N素重量损失率最高,达77%。造成两树种细根分解过程中N素释放程度不同的主要原因,可能是由于刺槐属固氮树种,细根中含N量高,C/N小,因此分解速度快,N素损失百分率高;杨树细根含N量低,C/N大,分解速度慢,N素损失百分率低。杨、槐细根混合分解N素损失率超过了刺槐细根单独分解的N素损失百分率,说明不仅是刺槐细根的供N作用促进了杨树细根的分解和养分释放过程,而且杨、槐细根在混合条件下有相互促进分解的作用。杨、槐细根分解过程中P素释放情况与N素类似,但杨树细根与刺槐细根单独分解时P素重量损失百分率的差别不如N素那么大。P素损失百分率:杨树细根在刺槐纯林下分解为62.5%,刺槐细根单独分解为64.5%~70.0%,二者接近;杨、槐细根混合分解介于两树种细根单独分解之间。可见与刺槐细根混合可促进杨树细根的P素养分释放速度,这对于改善混交林中刺槐的P素情况是有利的,而刺槐P素养分状况的改善又能加强其固氮能力。进而改善杨树的N素状况。于是混交林中杨树与刺槐之间在N、P养分方面就形成了一种互补关系。
|
|

细根单独分解和混合分解时都以K素养分的重量损失百分率最大,达90%以上,而且5种分解条件下差别不大,这可能是因为K素在植物体中主要以离子形态存在,极易被雨水和土壤水分淋溶而丢失。
Ca、Mg元素的释放规律相似,都表现为杨树细根单独分解的重量损失百分率低于刺槐细根单独分解,而杨、槐细极混合分解的损失百分率介于两者之间;元素的损失百分率Mg比Ca高,但都低N、P、K,这可能是因为Ca,Mg元素在植物组织中常常以较难溶解的化合物如硫酸盐、磷酸盐等形式被固定,迁移性差,不易被雨水淋失。杨树细根在刺槐纯林下分解比在其自身纯林下分解的Ca、Mg损失百分率差别不大,可见刺槐对于杨树细根分解过程中Ca,Mg元素的释放和归还的促进作用。
3.2 不同林分的养分归还目前国内对森林生态系统细根周转的研究尚不多见,对细根分解及其养分归还的研究更是少见报道(李凌浩,1998;廖利平等,1999)。本文得出了不同林分土壤通过细根死亡得到的养分归还量(见表 3)。
|
|
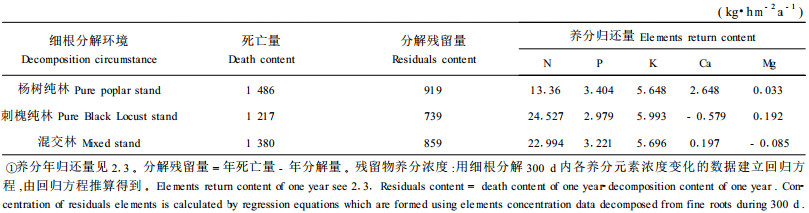
从表 3看出,不同林分的细根N素年归还量大小顺序为;刺槐纯林、混交林、杨树纯林,而且不同林分相差很大,刺槐纯林、混交林细根的N素年归还量分别为杨树纯林的18.4倍和17.2倍,可见通过与固氮树种刺槐形成混交林可大大提高林分细根的N素归还量,有利于杨树的生长。
不同林分细根的P素年归还量大小顺序为;杨树纯林、混交林、刺槐纯林、3种林分的P素年归还量差别不大,前两者分别只比第三者高14.3%和8.1%,但是混交林细根土壤的P素归还量较刺槐纯林有所提高,这对混交林中刺槐的生长和固氮作用是有好处的,良好的P素营养状况可促进固氮树种的固氮作用。
K素的年归还量变化规律与N素相同;刺槐纯林细根的K素年归还量最大,混交林次之,杨树纯林最小,但三者的差异非常小。3种林分中细根Ca元素的年归还量以杨树纯林最大,为混交林的13.4倍,而刺槐纯林的Ca元素不但不向土壤归还,反而向死细根积累(表现为年归还量为负值),其原因尚不清楚。3种林分中细根的Mg元素年归还量为:刺槐纯林>杨树纯林,前者为后者的5.8倍,混交林细根的Mg元素年归还量为负值。这说明杨槐混交林的细根在分解过程中出现了Mg元素的富积现象,其原因有待研究。
上述研究表明,细根分解过程中不同元素含量的变化情况不一样,N、P、K元素在3种林分中都表现为死细根向土壤的释放;Ca、Mg元素则在刺槐纯林和混交林的细根分解中表现为养分积累。不同林分在细根分解过程中各养分元素含量的变化有较大的差异:混交林细根的N、K、Mg元素年归还量大于杨树纯林而小于刺槐纯林;P、Ca元素的年归还量小于杨树纯林而大于刺槐纯林。总的来看,杨槐混交林细根的N素归还量大于杨树纯林,P素归还量大于刺槐纯林。从细根周转和分解过程中养分年归还量的角度看,杨槐混交林的N素和P素养分状况分别比杨树纯林和刺槐纯林有明显的改善,这对于协调混交林中杨树和刺槐两树种之间的N、P养分关系有着重要的意义。
4 结论树木细根是森林生态系统地下部分“凋落物"的主要组成部分,细根的养分含量比地上的枯枝落叶高,分解速度又比枯枝落叶快,因此在研究森林生态系统凋落物积累、分解、养分归还时,必须考虑细根的作用。
杨树细根在分解前期,N、Ca、Mg元素的浓度逐渐升高,P、K元素浓度则逐渐降低;刺槐细根分解过程中N、P、K元素浓度逐渐降低,Ca、Mg元素浓度逐渐升高;杨,槐细根混合分解各元素浓度变化介于两树种细根单独分解之间。
细根的N、K、Mg年归还量以刺槐纯林最大,混交林次之,杨树纯林最小。混交林中N素归还量的增加有利于改善杨树的N素状况。P、Ca元素的归还量大小顺序为:杨树纯林、混交林、刺槐纯林;杨树细根的P素归还量大于刺槐细根,这对于混交林中刺槐的生长有积极的意义和重要的作用。
杨槐栽后促进了杨树细根的周转,分解,养分释放与归还过程,尤其是P素养分归还量的增加可促进刺槐的固氮作用;刺槐细根的周转和分解使混交林的N素归还量比杨树纯林有明显的增加,从而改善了混交林土壤的N素养分状况,有利于杨树生长。混交林杨树与刺槐在N、P养分方面形成协调互补的关系。
沈国舫, 贾黎明, 翟明普. 1998. 沙地杨树刺槐人工混交林的改良土壤功能及养分互补关系. 林业科学, 34(5): 12-20. DOI:10.3321/j.issn:1001-7488.1998.05.002 |
单建平, 陶大立, 王淼, 等. 1993. 长白山阔叶红松林细根周转的研究. 应用生态学报, 4(3): 241-245. DOI:10.3321/j.issn:1001-9332.1993.03.008 |
贾黎明, 翟明普, 智信, 等. 1997. 杨树刺槐混交林生长状况及生产力. 北京: 中国林业出版社, 4.
|
廖利平, 杨跃军, 汪思龙, 等. 1999. 杉木火力楠纯林及其混交林细根分布、分解与养分归还. 生态学报, 19(3): 342-346. DOI:10.3321/j.issn:1000-0933.1999.03.009 |
廖利平, 高洪, 于小军, 等. 2000. 人工混交林中杉木、桤木和刺楸细根养分迁移的初步研究. 应用生态学报, 11(2): 161-164. DOI:10.3321/j.issn:1001-9332.2000.02.001 |
翟明普, 贾黎明, 沈国舫. 1997. 杨树刺槐混交林及树种间作用机制的研究. 北京: 中国林业出版社, 3-10.
|
翟明普, 蒋三乃, 贾黎明. 2002. 杨树刺槐混交林细根动态. 北京林业大学学报, 24(5-6): 39-44. |
Finer L, Christian M and Louis D G. 1997. Fine-root dynamics in mixed boreal conifer-broad-leafed forest stands at different successional stages after fire. Can J For Res, 27: 304-314. DOI:10.1139/x96-170 |
Hendrick R L & Pregitzer K S. 1992. The demography of fine root in a northern hardwood forest. Ecology, 73: 1094-1104. DOI:10.2307/1940183 |
McClaugherty C A, Aber J D, Melillo J M. 1982. The role of fine root in the organic matter and nitrogen budgets of two forested ecosystems. Ecology, 63: 1481-1490. DOI:10.2307/1938874 |
Nadelhoffer K J & Raich J W. 1992. Fine root production estimates and below-ground carbon allocation in forest ecosystems. Ecology, 73: 1139-1147. DOI:10.2307/1940664 |
Robinson C H, Kirkham J B, Littlewood R. 1999. Decomposition of root mixtures from high arctic plants: a microcosm study. Soil Biol Biochem, 31: 1101-1108. DOI:10.1016/S0038-0717(99)00028-0 |
Roy S & Singh J S. 1995. Seasonal and spatial dynamics of plant-available N and P pools and N-mineralization in relation to fine roots in a dry tropical forest habitat. Soil Biol Biochem, 27: 33-40. DOI:10.1016/0038-0717(94)00138-Q |
Ruess R W, Cleve K, Yarie J, et al. 1996. Contribution of fine root production and turnover to the carbon and nitrogen cycling in taiga forests of the Alaskan interior. Can J For Res, 26: 1326-1336. DOI:10.1139/x26-148 |
Thorup-Kristensen K & Riki B. 1998. Temporal and spatial root development of cauliflower (Brassica oleracea var. bootrytis). Plant and Soil, 201: 37-47. DOI:10.1023/A:1004393417695 |
Vogt K A, Gerier C C, Vogt D J. 1986. Production, turnover and nutrient dynamics of above-and below-ground detritus of world forests. Adv Ecol Res, 15: 303-377. DOI:10.1016/S0065-2504(08)60122-1 |