 2004, Vol. 40
2004, Vol. 40文章信息
- 陈光水, 何宗明, 谢锦升, 杨玉盛, 蒋宗垲.
- Chen Guangshui, He Zongming, Xie Jinsheng, Yang Yusheng, Jiang Zhongkai.
- 福建柏和杉木人工林细根生产力、分布及周转的比较
- Comparision on Fine Root Production, Distribution and Turnover Between Plantations of Fokienia Hodginsii and Cunninghamia Lanceolata
- 林业科学, 2004, 40(4): 15-21.
- Scientia Silvae Sinicae, 2004, 40(4): 15-21.
-
文章历史
- 收稿日期:2002-07-25
-
作者相关文章


2. 福建农林大学林学院 南平 353001;
3. 福建农林大学莘口教学林场 三明 365002
2. Forestry College of Fujian Agriculture and Forestry University Nanping 353001;
3. Xinkou Experimental Forest Farm of Fujian Agriculture and Forestry University Sanming 365002
林木细根具有巨大的吸收表面积,是林木吸收水分和养分的主要器官,同时因其生长和周转迅速,对森林生态系统物质循环和能量流动起着十分重要的作用(Vogt et al., 1986; Ruess et al., 1996; Gordon et al., 2000; Chen et al., 2002) 。虽然细根占林木根系总生物量的不足30%,但其年净生产力却占森林总净生产力的30% ~80%;每年通过枯死细根向土壤归还的碳、养分和能量甚至超过地上凋落物(Vogt et al., 1986; Santantonio et al., 1987; Grier et al., 1981; McClaugherty et al., 1982; Burke et al., 1994; Gordon et al., 2000) 。
虽然对林木根系的研究早在18世纪初就已开始,但直到Harris(1977) 首次证明了细根的生产量与其生物量不成比例之后,细根生长、死亡、寿命、生物量、生产量、周转量、分布、季节动态及细根对森林生态系统C和养分循环的贡献等研究才迅速增加,并逐渐成为国际性研究热点之一(Persson, 1983; McClaugherty et al., 1982; Vogt et al., 1986; Santantonio et al., 1987) 。20世纪90年代初,微根管(minirhizostron) 技术开始在林木细根研究中得到广泛应用,推动细根研究进一步深入(Hendrick et al., 1993; Sovan et al., 1995; Arunachalam, 1996; Zogg et al., 1996) 。最近,大量的研究者通过模拟大气CO2浓度、臭氧浓度、气温升高等,研究细根对全球变化的响应(King et al., 1996;Janssens et al., 1998; Retzlaff et al., 1996; Pregitzer et al., 2000) 。我国与国外有关林木细根的同类研究水平相差甚远,早期国内采用全挖法研究根系生物量(张其水等,1991) ,后应用全挖法(粗根) 配合土柱法(细根) 研究了根系的生物量(杨玉盛等,1991;李振问等,1993) 。90年代中后期,国内研究人员开始采用土芯法(soil core) 和内生长土芯法(ingrow core) 研究细根的生物量、生产力、分布和细根的养分归还等(单建平等,1993;廖利平等,1995;李凌浩等,1998;温达志等,1999;杨玉盛等, 2001, 2002;陈光水等,2001) 。本文从细根生产力、分布及周转角度比较福建柏林与杉木林生态学差异。
1 试验地概况试验地位于三明莘口教学林场小湖工区(北纬26°11′30″,东经117°26′00″) ,福建柏(Fokienia hodginsii) 和杉木(Cunninghamia lanceolata) 人工林前身均为格氏栲(Castanopsis kawakamii) 、米槠(Castanopsis carlesii) 等为主的天然林,1966年经皆伐劈草炼山,穴状整地,1967年初用福建柏和杉木实生苗营造人工纯林,试验地气候条件、林分抚育措施、标准地概况、土壤理化性质等详见文献(杨玉盛等,2004a) 。
2 研究方法 2.1 细根生物量在福建柏林和杉木林中各设立3块20 m×20 m固定标准地,从1999年1月至2001年11月隔月于月底用内径6.8 cm的土钻在各林分标准地的上中下部随机钻取土芯10个,每个林分共30个,深度为1 m。取出土芯按0~10、10~20、20~30、30~40、40~50、50~60、60~70、70~80、80~90及90~100 cm分割土芯,分别用流动水浸泡、漂洗、过筛,拣出根系,用放大镜、剪刀、镊子等工具分别各层次分出各目的树种根,捡出直径<2 mm的细根,并根据根系外形、颜色、弹性、根皮与中柱分离的难易程度来区分活死根,对目的树种细根进一步细分成3个径级(1~2 mm、0.5~1 mm、<0.5 mm) ,将各根样品置于105℃烘箱中烘干至恒重后称重,并按以下公式计算细根生物量(McClaugherty et al., 1982) :细根现存量(g·m-2) =平均每根土芯细根干重(g) ×/〔π(6.8 cm/2) 2×(m2/10 4cm2) 〕。
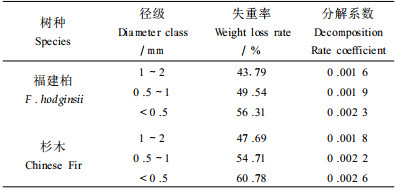
2.2 细根分解试验随机收集福建柏和杉木林表层土(0~20 cm) 的活根与死根混合样品,分成3个径级自然风干后,称取5 g(每个树种、每个径级) 样品装入18 cm×18 cm、孔径为0.25 mm的尼龙网袋中(根据样品含水量换算为干重) ,埋入各自标准地中部10 cm深处,每个径级共80袋,并于放置0、30、60、90、150、210、270、330、390、510、630、750 d后随机抽取各径级样品各6袋,用于失重和化学成分分析。根据X/X0=exp(-kt) (其中,X0为分解初始干重,X为分解t d后的残留物干重,k为分解系数) ,建立细根不同径级指数方程,求得分解系数k。
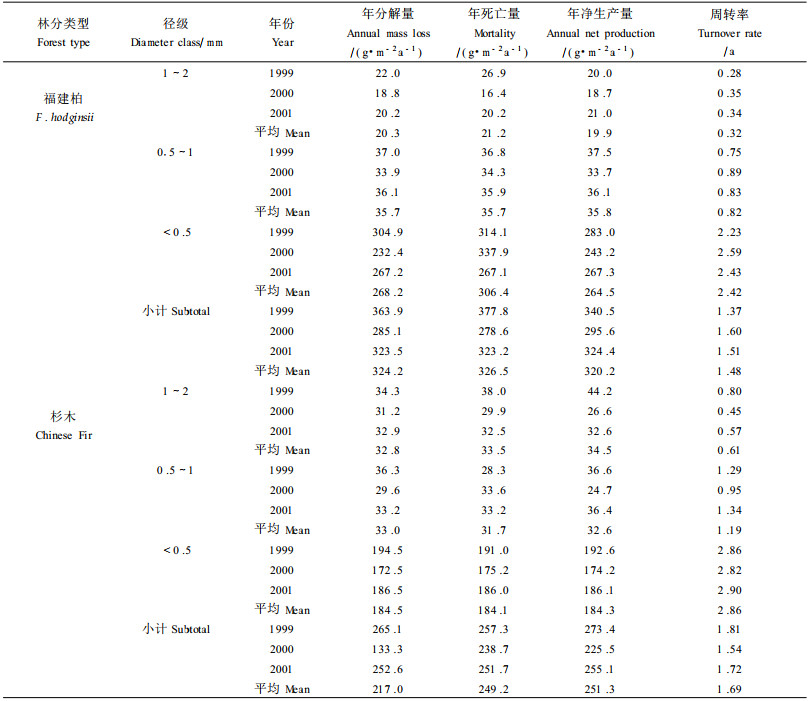
2.3 细根净生产力和周转细根年净生产量、分解量和死亡量用改进的分室通量模型计算(Santantonio et al., 1987; Kurz et al., 1987) :LFRt=LFRt-l+Pt-Mt; DFRt=DRTt-l+Mt-Dt; Dt=(DFRt-l+Mt) DRt; T=P/Y, 式中:LFR:活细根生物量,DFR:死细根生物量,P:年净生产量,M:年死亡量,D:年发解量,DR:分解速率,T:周转速率,Y:平均活细根生物量,t为时间间隔。
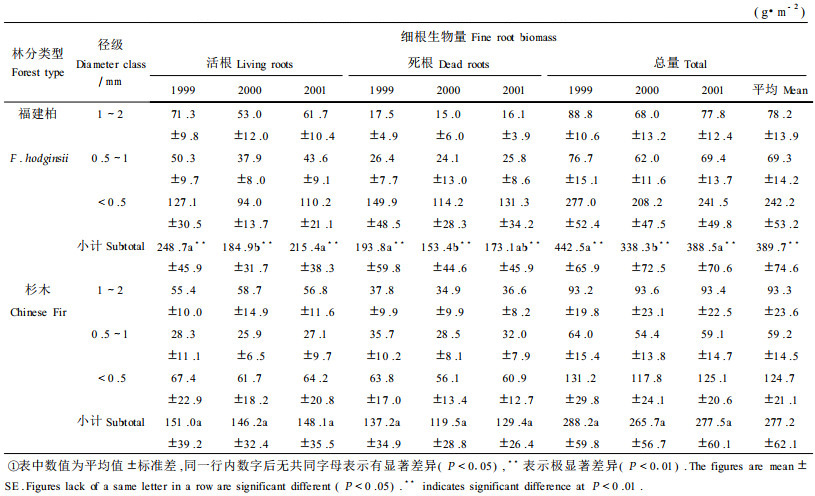
3 结果分析 3.1 细根生物量及分布福建柏林1999—2001年细根生物量1999年与2000年间存在显著差异(P<0.05) ,而杉木林3?a细根生物量两两之间差异均不显著(P>0.05) ;福建柏与同年度杉木细根生物量间均存在极显著差异(P<0.01) (表 1) 。福建柏年均细根生物量达389.7 g·m-2,是杉木林(277.2 g·m-2) 的1.41倍;活细根年均生物量达216.3 g·m-2,是杉木林(148.4 g·m-2) 的1.46倍(P<0.05) 。虽然<0.5 mm细根是两林分细根的主体,但福建柏林<0.5 mm细根所占比例(61.2%) 却比杉木林(45.0%) 的高出16.2%,而<0.5 mm细根生物量(242.2 g·m-2) 则是杉木林(124.7 g·m-2) 的1.94倍(P<0.05) 。福建柏1~2 mm、0.5~1 mm和<0.5 mm径级细根生物量中死细根所占比例分别为20.7%、36.7%和54.4%,而杉木的则分别为39.0%、54.2%和48.3% (表 2) 。福建柏林总细根生物量中死细根生物量所占比例(44.5%) 则与杉木林(46.4%) 的相似。
|
|
|
|
福建柏和杉木的总细根密度在0~10 cm土层差异最大,福建柏和杉木的分别达144.2 g·m -2和70.2g·m-2,前者是后者的2.1倍,10~20 cm土层福建柏总细根密度(78.6 g·m-2) 仍比杉木的(58.4 g·m-2) 高出34.6%;而在20 cm深度以下福建柏和杉木细根密度较为接近(图 1) 。0~10 cm土层福建柏细根总量占福建柏细根总量的37.0%,比杉木的(25.3%) 高出11.7%。

|
图 1 不同林分细根垂直分布 Fig. 1 Vertical distribution of fine root in different plantations ●福建柏细根总量Total amount of F.hodginsii fine roots ○福建柏活细根Living fine roots of F.hodginsii ▲杉木细根总量Total amount of Chinese Fir fine roots △杉木活细根Living fine roots of Chinese Fir |
3年中福建柏和杉木活细根生物量均有明显的季节差异(P<0.05) (图 2) 。福建柏林和杉木林活细根生物量每年均在3月出现峰值,但福建柏2000年3月峰值显著低于1999年和2001年(P<0.01) ,而杉木的则无显著差异(P>0.05) ;杉木林活细根生物量还在9月出现第2个峰值,而福建柏的则不明显。两种林分活细根生物量最低值均分别出现在5月(7月) 和11月(1月) (图 2) 。两种林分死细根生物量最大值均出现在5月(7月) ;死细根生物量最低值出现时间福建柏林为7月(9月) 和1月,而杉木的则为3月和9月(图 2) 。

|
图 2 不同林分细根生物量季节动态 Fig. 2 Seasonal dynamics of fine root biomass in different plantations ●活细根Living fine roots ○死细根Dead fine roots |
福建柏和杉木不同径级细根的第1年失重率及分解系数(k) 随径级的减小而增大,且福建柏各径级细根第1年分解失重率及k值均低于相应径级杉木细根(表 2) 。福建柏林细根年死亡量为326.5 g·m-2a-1,是杉木(249.2 g·m-2a-1) 的1.31倍;细根年净生产量为320.2 g·m-2a-1,是杉木(251.3 g·m-2 a-1) 的1.27倍(表 3) 。福建柏<0.5 mm径级细根占总细根年死亡量和年净生产量的比例分别为93.84%和82.60%,均分别高于杉木的(73.88%和73.34%) 。福建柏细根3年平均周转速率为1.48 a-1,比杉木(1.69 a-1) 低(表 3) 。
|
|
Vogt等(1996) 综述世界上有关亚热带森林细根报道后指出其生物量介于1.1~5.8 t·hm -2之间,本研究两林分乔木细根生物量落入其中(表 1) 。与国内同处亚热带的其它森林类型相比,福建柏林细根生物量高于湖南会同的杉木纯林和杉木火力楠(Michelia macclurei) 混交林,与湖南会同火力楠纯林和本试验地27年生杉木林的相似,而低于中亚热带武夷山的甜槠(Castanopsis eyeri) 林和南亚热带鼎湖山季风常绿阔叶林和针阔叶混交林(廖利平等,1995;杨玉盛等,2001;李凌浩等,1998;温达志等,1999) 。福建柏和杉木细根生物量分别占其乔木层生物量的1.70%和1.18%,这与世界上许多森林类型的研究结果相似(Grier et al., 1981; Arthur et al., 1992; Hendrick et al., 1993; Ruess et al., 1996; 单建平等,1993;杨玉盛等,2002) 。本研究中福建柏和杉木林<0.5 mm细根是细根的主体,这与前人的研究结果相似(Hendrick et al., 1993; McClaughterty et al., 1982;杨玉盛等,2002) 。对福建柏和杉木标准木采用全挖法获取的<2 mm细根分别为123.8和55.9 g·m-2,仅占土芯法的31.77%和20.17%。因此应该采用全挖法确定>2 mm粗根,及土芯法确定<2 mm细根,才能提高根系生物量测定精度(杨玉盛等,1991;2001) 。
本研究中福建柏细根生物量(特别是活细根量) 、<0.5 mm细根数量及其占细根总量比例比杉木的高,以及福建柏细根高度密集于0~20 cm(特别是0~10 cm) 土层(图 1) ,体现福建柏和杉木树种生物学特性差异。由于福建柏凋落物数量大、N、P养分浓度高、分解速率快(杨玉盛等,2004a; 2004b) 及表层土壤肥力比杉木林的高,亦可能加大其表层土壤细根密度。另则福建柏细根密集于表层土壤,能迅速吸收枯枝落叶层分解释放的养分及降雨淋溶中的养分,加上细根对表土的穿插切割作用及伴随细根枯死归还有机质、养分,反过来促进表层土壤肥力进一步提高。
4.2 细根生物量动态细根生物量一年中常出现1或2个峰值,或季节变化不明显;峰值出现在春季展叶期前后、晚夏或秋季等,受树种特性及外界环境条件(如降水量、土温、养分有效性等) 综合影响,细根生物量季节动态仍会波动(McClaugherty et al., 1982; Grier et al., 1981; Santantonio et al., 1987; Hendrick et al., 1993; 1996;Mallonen et al., 1998; Fahey et al., 1994; 单建平等,1993;温达志等,1999;杨玉盛等,2001) 。本研究中福建柏和杉木活细根量在3月出现峰值与亚热带此时地温回升、土壤湿度增加有关(图 2) 。9月(俗称“小阳春”) ,杉木活细根生物量与地上部分树高、胸径生长相应出现第2个峰值,而福建柏则不明显(图 2) 。5月(7月) 两种林分死细根生物量均出现峰值可能与此时活细根大量死亡有关。本研究中细根生物量的季节变化与鼎湖山、长白山等地的研究结果不尽相同(单建平等,1993;温达志等,1999) ,与研究地自然条件及树种特性等不同有关。
对福建柏和杉木活细根生物量与月降雨量、月均气温等气候因子的相关分析表明,福建柏活细根量与当月降雨量呈显著的正相关,而杉木活细根生物量则均与气候因子无显著相关;且福建柏细根生物量年际间差异大于杉木的(表 1) 。表明福建柏细根生长除受自身生长节律影响外,还受气候条件波动的影响。这与福建柏凋落物凋落节律受降雨与温度影响相似(杨玉盛等,2004a) 。表明福建柏细根和树叶生长发育比杉木更易受气候波动影响。这与该地属杉木中心产区,而非福建柏最适宜生长区有关(其最适宜区在中亚热带南部及其以南地区) (章浩白,1993) 。
4.3 细根净生产力和周转福建柏林和杉木林的细根净生产力均在已报道的世界森林类型细根净生产力范围内(1.4~11.5 t·hm-2a-1) (Santantonio et al., 1987) 。与亚热带森林类型相比,本研究中福建柏年净生产力高于会同11年生杉木纯林、杉木火力楠混交林及南亚热带鼎湖山针阔叶混交林,低于会同11 a生火力楠纯林、鼎湖山季风常绿阔叶林及武夷山76 a生甜槠林,而与本地27 a生杉木纯林和杉木观光木(Tsoongiodendron odorum) 混交林相近(廖利平等,1995;李凌浩等,1998;温达志等,1999;杨玉盛等,2001) 。本研究中福建柏和杉木细根生物量仅占其乔木层生物量的1%~2%,但其细根净生产力却均占其乔木层总净生产力(16.143) 和13.083 t·hm-2a-1,) 的19.84%和19.21%,低于该地的27 a生杉木观光木混交林(22.9%) 和杉木纯林(20.9%) (杨玉盛等,2001) ,但高于南亚热带鼎湖山季风常绿阔林和针阔中混交林细根(<5 mm) 所占的比例(16.8%和17.4%) (温达志等,1999) 。与世界其它气候带森林相比,亚热带森林细根的净生产力所占比例普遍较低,其原因有待于进一步探讨。细根枯死是森林生态系统地下部分“枯落物”的主要组成部分(Vogt et al., 1986) ,本研究中福建柏和杉木细根年均死亡量分别是地上部分凋落物量(669.6和488.6 g·m-2) 的48.74%和51.00%(杨玉盛等,2004a) ,落于已报道的范围内(Vogt et al., 1986; Arthur et al., 1992; Hendrick et al., 1993, 温达志等,1999;杨玉盛等,2001) 。
Gill等(2000) 综述世界各地的研究表明,世界各森林类型细根周转速率范围在0.019~2.644 a-1之间,平均周转速率为0.56 a-1,细根周转速率与年平均气温间存在弱的正相关,并随纬度的增大而减小。本研究中福建柏和杉木细根年周转速率均落入已报道的细根周转速率范围,且均高于世界森林细根平均周转速率(表 3) 。与亚热带森林类型相比,本研究中福建柏细根周转速率高于鼎湖山的季风常绿阔叶林和针阔叶混交林、武夷山的甜槠林、会同的11年生杉木纯林、火力楠纯林及杉木火力楠混交林及本试验地27年生杉木林(廖利平等,1995;李凌浩等,1998;温达志等,1999;杨玉盛等,2001) 。
陈光水, 林瑞余, 卢镜铭, 等. 2001. 杉木观光木混交林细根灰分含量和热值动态. 福建林学院学报, 24(4): 332-335. DOI:10.3969/j.issn.1001-389X.2001.04.012 |
单建平, 陶大立, 王森, 等. 1993. 长白山阔叶红松林细根周转的研究. 应用生态学报, 4(4): 241-245. |
李凌浩, 林鹏, 邢雪荣. 1998. 武夷山甜槠林细根生物量和生长量研究. 应用生态学报, 9(4): 337-340. DOI:10.3321/j.issn:1001-9332.1998.04.001 |
李振问, 杨玉盛. 1993. 杉木火力楠混交林根系的研究. 生态学杂志, 12(1): 20-24. DOI:10.3321/j.issn:1000-4890.1993.01.008 |
廖利平, 陈楚莹, 张家武, 等. 1995. 杉木、火力楠纯林及混交林细根周转的研究. 应用生态学报, 6(1): 7-10. DOI:10.3321/j.issn:1001-9332.1995.01.005 |
温达志, 魏平, 孔国辉, 等. 1999. 鼎湖山南亚热带森林细根生产力与周转. 植物生态学报, 23(4): 361-369. DOI:10.3321/j.issn:1005-264X.1999.04.009 |
杨玉盛, 陈光水, 何宗明, 等. 2002. 杉木观光木混交林和杉木纯林群落细根生产力、分布及养分归还(英文) . 应用与环境生物学报, 8(3): 223-233. DOI:10.3321/j.issn:1006-687X.2002.03.001 |
杨玉盛, 陈光水, 何宗明, 等. 2001. 杉木观光木混交林群落细根净生产力及周转. 林业科学, 37(专刊1): 35-41. |
杨玉盛, 陈银秀, 何宗明, 等. 2004a. 福建柏和杉木人工林凋落物性质的比较. 林业科学, 40(1): 2-10. |
杨玉盛, 郭剑芬, 陈银秀, 等. 2004b. 福建柏和杉木人工林凋落物分解及养分动态的比较. 林业科学, 40(3): 19-25. |
杨玉盛, 林先富, 俞新妥, 等. 1991. 杉木套种山苍子模式的结构与生物量初步研究. 福建林学院学报, 11(4): 341-348. |
张其水, 俞新妥. 1991. 连栽杉木林的根系研究. 植物生态与地植物学报, 15: 374-379. |
章浩白. 1993. 福建森林. 北京: 中国林业出版社.
|
Arthur M A, Fahey T J. 1992. Biomass and nutrients in an Engelmann spruce-subalpine fir forest in north central Colorado:pools, annual production, and internal cycling. Can J For Res, 22: 315-325. DOI:10.1139/x92-041 |
Arunachalam A. 1996. Fine root decomposition and nutrient mineralization patterns in a subtropical humid forest following tree cutting. For Ecol Manag, 86(1/3): 141-150. |
Burke M K, Raynal D J. 1994. Fine root growth phenology, production, and turnover in a northern hardwood forest ecosystem. Plant and Soil, 162: 135-146. DOI:10.1007/BF01416099 |
Chen H, Harmon M E, Sexton J, et al. 2002. Fine-root decomposition and N dynamics in coniferous forests of the Pacific Northwest, U S A. Can J For Res, 32: 320-331. DOI:10.1139/x01-202 |
Fahey T J, Hughes J W. 1994. Fine root dynamics in a northern hardwood forest ecosystem.Hubbard Brook Experimental Forest. NH.Ecology, 82: 533-548. |
Gill R A, Jackson R B. 2000. Global patterns of root turnover for terrestrial ecosystems. New Phytol, 147: 13-31. DOI:10.1046/j.1469-8137.2000.00681.x |
Gordon W S, Jackson R B. 2000. Nutrient concentrations in fine roots. Ecology, 81(1): 275-280. DOI:10.1890/0012-9658(2000)081[0275:NCIFR]2.0.CO;2 |
Grier C C, Vogt K A, Keyes M R, et al. 1981. Biomass distribution above and below-ground in young and mature Abies amabilis zone ecosystems in the Washington Cacades. Can J For Res, 11: 155-167. DOI:10.1139/x81-021 |
Harris W F. 1977. Comparison of below-ground biomass in natural deciduous forests and loblolly pine plantation. Pedobiologia, 17: 369-381. |
Hendrick R L, Pregitzer K S. 1996. Temporal and depth-related patterns of fine root dynamics in northern hardwood forest. Ecology, 84: 167-176. DOI:10.2307/2261352 |
Hendrick R L, Pregitzer K S. 1993. The dynamics of fine root length, biomass, and nitrogen content in two northern hardwood ecosystems. Can J For Res, 23: 2507-2520. DOI:10.1139/x93-312 |
Janssens I A, Crookshanks M, Ceulemans R. 1998. Elevated atmospheric CO2 increases fine root production, respiration, rhizosphere respiration and soil CO2 efflux in Scots pine seedlings. Glob Change Biol, 4(8): 871-878. DOI:10.1046/j.1365-2486.1998.00199.x |
King J S, Thomas R B, Strain B R. 1996. Growth and carbon accumulation in root systems ofPinus taeda and Pinus ponderosa seedlings as affected by varying CO2, temperature and nitrogen. Tree Physiol, 16(7): 635-642. DOI:10.1093/treephys/16.7.635 |
Kurz W A, Kimmins J P. 1987. Analysis of some sources of error in methods used to determine fine root production in forest ecosystems:a simulation approach. Can J For Res, 17: 909-912. DOI:10.1139/x87-142 |
Mallonen K, Helmisaari H S. 1998. Seasonal and yearly variations of fine-root biomass and necromass in a Scots pine (Pinus sylvestris L.) stand. For Ecol Manag, 102: 283-290. DOI:10.1016/S0378-1127(97)00169-2 |
McClaugherty C A, Aber J D. 1982. The role of fine roots in the organic matter and nitrogen budgets of two forested ecosystems. Ecology, 63: 1481-1490. DOI:10.2307/1938874 |
Persson H. 1983. The distribution and productivity of fine roots in boreal forests. Plant and Soil, 71: 87-101. DOI:10.1007/BF02182644 |
Pregitzer K S, Zak D R, Maziasz J. 2000. Interactive effects of atmospheric CO2 and soil-N availability on fine roots of Populus tremuloides. Ecol Appli, 10(1): 18-33. |
Retzlaff W A, Weinstein D A, Laurence J A. 1996. Simulated root dynamics of a 160-year-old sugar maple (Acer saccharum Marsh.) tree with and without ozone exposure using the TREGRO model. Dynamics of physiological processes in woody roots.Tree Physiol, 16(11/12): 915-921. |
Ruess R W, Cleve K V, Yarie J, et al. 1996. Contributions of fine root production and turnover to the carbon and nitrogen cycling in taiga forests of the Alaskan interior. Can J For Res, 26: 1326-1336. DOI:10.1139/x26-148 |
Santantonio D, Grace J C. 1987. Estimating fine-root production and turnover from biomass and decomposition data:a compartment-flow model. Can J For Res, 17: 900-908. DOI:10.1139/x87-141 |
Sovan R, Singh J S. 1995. Seasonal and spatial dynamics of plant-available N and P pools and N-mineralization in relation to fine roots in a dry tropical forest habitat. Soil Biol Biochem, 27: 1. DOI:10.1016/0038-0717(94)00166-X |
Vogt K A, Grier C C, Vogt D J. 1986. Production, turnover and nutrient dynamics of above-and below-ground detritus of world forests. Adv Ecol Res, 15: 303-377. DOI:10.1016/S0065-2504(08)60122-1 |
Vogt K A, Vogt D J, Palmiotto P A, Boon P. 1996. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant and soil, 187: 159-219. |
Zogg G P, Zak D R, Burton A J. 1996. Fine root respiration in northern hardwood forests in relation to temperature and nitrogen availability. Tree Physiol, 16(8): 719-725. DOI:10.1093/treephys/16.8.719 |