 2004, Vol. 40
2004, Vol. 40文章信息
- 陈鹏, 何凤仁, 钱伯林, 韦军, 王莉.
- Chen Peng, He Fengren, Qian Bolin, Wei Jun, Wang Li.
- 中国银杏的种核类型及其特征
- Seed Types and Their Relative Characteristics in Ginkgo biloba of China
- 林业科学, 2004, 40(3): 66-70.
- Scientia Silvae Sinicae, 2004, 40(3): 66-70.
-
文章历史
- 收稿日期:2002-10-17
-
作者相关文章


植物学上根据树冠形状和枝叶变化将银杏(Ginkgo biloba)分为塔型银杏、垂枝银杏、裂叶银杏、斑叶银杏、黄叶银杏和叶籽银杏等类型(陈鹏,1991;郭善基,1993)。银杏雌雄性别的形态与结构及其发生特点与鉴别已见较多的研究报道(王伏雄等,1983;Foster et al., 1983;Friedman,1987; Friedman et al., 1988;陈鹏,1991; 傅德志等,1993;陶俊等,1998;吉成均,1999)。我国银杏雌株资源丰富,长期以来以利用种核为主要栽培目的(何凤仁,1989;陈鹏等,1996;1997)。曾勉(1935)根据种核的形状和特征进行分类,将核用银杏分成梅核、佛手和马铃3种类型。目前,种核生产已成为我国银杏资源利用的主要特色和优势(陈鹏等,2000;陈鹏,2002)。由于银杏雌株既有实生变异,又有营养变异,因而种内变异较大,核用品种类型较多(何凤仁,1989;郭善基,1993;陈鹏等,1997;陈鹏,2002),需要提出适用于银杏种核的科学分类方法,并确定各个类型的种核形态特征。在具体划分银杏种核类型的基础上,进一步研究银杏种实的生长发育过程,明确各类型种核的有关成份含量和品质,为银杏核用品种的改良和利用提供依据。
1 材料与方法 1.1 研究过程20世纪50年代始,全面调查我国银杏主产区雌株资源的分布及数量,确定重点调查的代表雌株,采集其枝叶和种实制作标本,进行种实解剖观察,测定种核的各项形态指标,计算数量性状的平均数和变异系数,提出银杏种核的分类方法,确定不同类型的代表特征。对不同种核类型进行品质鉴评,分析核形与种仁中有关成份的含量(何凤仁,1989;陈鹏等,1994;1997;2000;Chen et al., 1999),明确类型间有关理化指标的差异。
1.2 种实的解剖观察选择3株生长结果正常的佛指品种树,从开花盛期至种实成熟期(4月20日—9月底),每7天每树随机取种实10粒(共30粒),FAA固定。用徒手切片法(早期)和石蜡切片法切片后用番红(1%)和固绿(0.5%)双重染色,镜检不同时期种实各部组织结构变化,绘制种实生长发育过程图。应用概率单位分析(莫惠栋,1992)描述种实的生长过程。
1.3 种核的测定分析1979年起,选取佛指、洞庭皇、七星果、马铃、梅核和龙眼等核用品种,每品种分别选取3株生长结果正常的成年大树作为代表雌株,种实成熟时,定点定树随机取10粒种实,每品种共取30粒种实分别用精确度为0.001的游标卡尺和天平测定种柄、种托、种核的长、宽、厚及单核重,选择比较稳定、可靠的种实特征进行分类。1990年10月《中国果树志·银杏卷》编委会召开全国银杏品质鉴评会,在参评的66个品种中每样本亦均随机取30粒种核,按照同样的分类方法测定各项指标,确定每一样本种核的所属类型。根据本研究的需要,每一类型随机选定5个品种,每一品种随机取5粒种核(共25粒),分别去壳后,置种仁于65~70℃的烘箱中烘至恒重。测定干物重后粉碎,用日本MRK公司的VS-KTP全自动凯氏定氮仪测定种仁中粗蛋白含量;用蒽酮比色法测定可溶性糖含量;用碘比色法分别测定直链淀粉和支链淀粉含量;用离子计及标准加入法测定氢氰酸含量(陈鹏等,1994;1997;2000;Chen et al., 1999)。
1.4 种核的品质鉴评根据《中国果树志·银杏卷》编委会确定的银杏种核品质鉴评指标及评分标准,在全国银杏品质鉴评会期间组织与会专家及代表对参评品种进行品质鉴评打分。各个类型均利用种核分析中所用的5个品种,对不同类型的鉴评得分进行方差分析和差异显著性测验(莫惠栋,1992)。
2 结果与分析 2.1 银杏种实的生长发育银杏种实体积增长的Probit分析表明,银杏雌株的开花后天数(x)与种实体积的概率单位(p)的关系为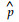

|
图 1 银杏种实的生长发育过程 Fig. 1 Seed growth and development course in G. biloba A种实的横切面Cross section of seed; B种实的纵切面Vertical section of seed. Ⅰ期:授粉期的胚珠(4月29日);Ⅱ期:珠被细胞迅速分裂,雌配子体游离核出现(5月13日);Ⅲ期:种实迅速膨大,雌配子体细胞壁形成(5月27日);Ⅳ期:种实迅速膨大,初生颈卵器形成(6月10日);Ⅴ期:中种皮细胞壁从合点端开始加厚(6月24日);Ⅵ期:种实缓慢增大,中种皮木质化(7月8日);Ⅶ期:种实缓慢增大,中种皮木质化(7月22日);Ⅷ期:种实成熟,种胚形成(9月30日)。 StageⅠ: Ovule at pollination stage thickened (29th April); StageⅡ: Integument cell divided rapidly, free nuclei of female gamete appeared(13th May); StageⅢ: Seed expanded rapidly, cell wall of female gamete formed(27th May); StageⅣ: Seed expanded rapidly, primary archegonium formed(10th June); StageⅤ: Cell wall of mid-seed-coat initially from the end of chalaza(24th June); StageⅥ: Seed expanding slowly, mid-seed-coat lignified initially(8th July); StageⅦ: Seed increasing slowly, mid-seed-coat lignifications(22nd July); StageⅧ: Seed ripened, embryo formed (30th September). |
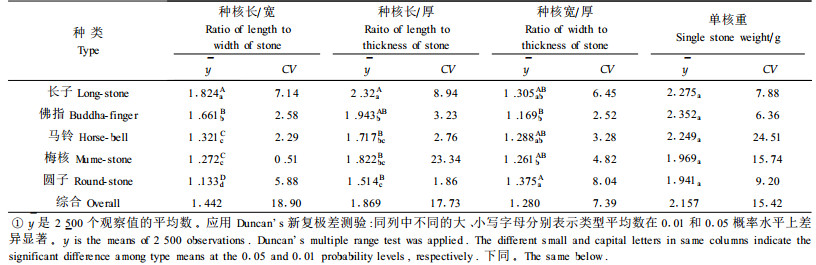
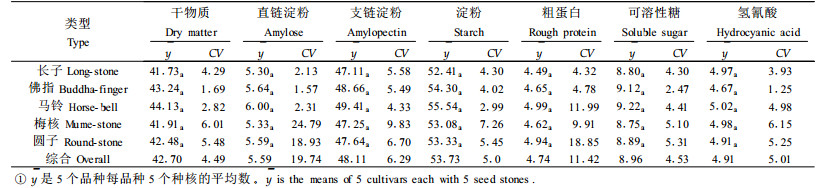
银杏种实遗传的稳定特征主要有珠托是否正中、种核的长宽比及形状、种核两侧有无明显的棱及是否呈翼状、种核先端圆钝或秃尖及有无孔迹突起等。综合种核形态特征,银杏雌株可分为长子、佛指、马铃、梅核、圆子5种类型(表 1)。种核的长/宽、长/厚、宽/厚在不同类型间有显著或极显著差异(表 2)。在长宽比上,长子类明显表现最大,其次为佛指类,再次为马铃、梅核和圆子类。由于种核的长宽比(x)与长厚比(y)间存在极显著正相关,前者每增加1个单位,后者平均增加1.065个单位(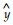
|
|

|
|
|
|
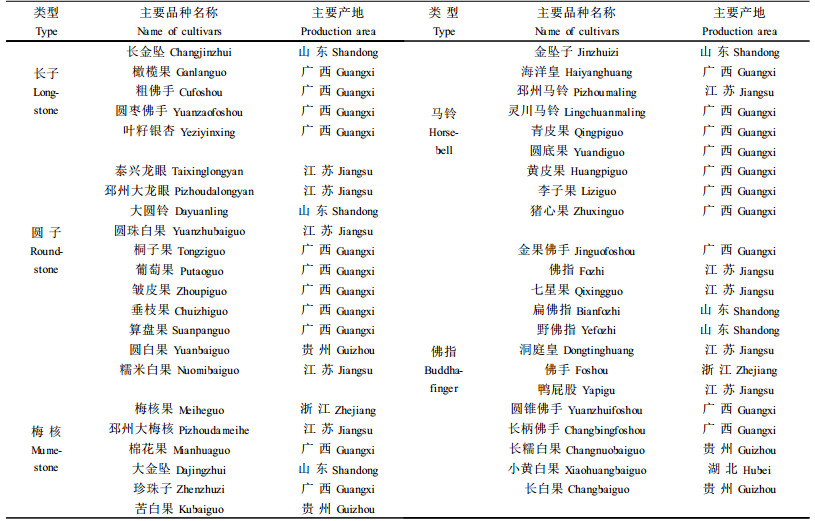
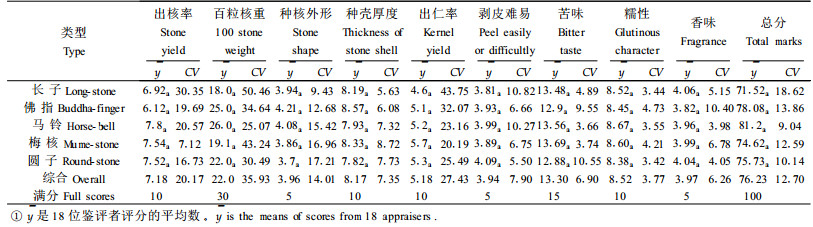
银杏种核的品质主要受干物质、淀粉、直链淀粉、支链淀粉,粗蛋白、可溶性糖及氢氰酸含量的影响。不同类型中,直链淀粉含量的差异最大,其次为粗蛋白含量,两者的变异系数分别为19.74%和11.42%;差异最小的为干物质含量,其变异系数仅为4.49%。但经方差分析,上述成份含量在不同类型间的差异均未达到显著水平(表 4)。但是,有些成分含量在类型内的植株间差异甚大。例如梅核和圆子类型的直链淀粉含量株间变异系数分别达到24.79%和18.93%;马铃和圆子类型的粗蛋白含量株间变异系数分别达到11.99%和18.85%。这说明单株选优仍将相当有效。单核重亦类似(表 2),其变异系数达到15.42%,但因株间变异较大,导致类型间差异不显著;而马铃和梅核的株间变异系数分别高达24.51%和15.74%, 显示了较大的选择潜力。以上情况与种核品质鉴评结果相一致(表 5)。例如百粒核重得分最高的有26.0分,高出最低的8.0分,类型间的变异系数达到35.9 3%。由于类型内的株间变异较大,最大的变异系数达到50.46%(长子),因而类型间百粒核重得分差异仍不显著。各种类型种核品质有关性状鉴评得分的变异大小依次为百粒核重、出仁率、出核率、种核外形、剥皮难易、种壳厚度、苦味、香味和糯性。由于类型内株间变异较大,类型间差异均未达到显著水平。以上说明品质性状的优劣在类型间呈随机分布,即各类型内均有优劣不同的品种,以致类型间没有明显的优劣差异。
|
|
|
|
由于银杏雌株主要生产种核,而作为生殖器官的种核具有稳定的遗传性状,因而根据种核的形态特征对银杏雌株品种进行分类具有科学性、可靠性和实用性。实践上,根据种核形状和长、宽变化确定雌株品种的类型(表 2),也简便易行。对于种核形状相似的品种,还可根据种实和种核的其它明显特征进一步鉴别(Davidson,1994)。例如佛指类型中有佛指和佛手2个品种,种核都呈长卵圆形,但可根据种实、种核的顶端有无秃尖及孔迹是否突起,将2品种分开(何凤仁,1989)。为了进一步明确品种类型间的遗传演变与谱系关系,当然还可采用同工酶及分子标记分析等方法,作深入研究。
3.2 银杏雌株品种的遗传改良本研究中,单核重及有关品质成分的测定结果与种核品质鉴评结果基本一致,即由于类型的植株间的变异较大,类型间的平均差异均不显著。这说明银杏遗传改良的重点仍应是在类型内选择优树,并通过嫁接进行繁殖,同时在各类型中都有可能选出和繁殖更好的优树。这为银杏雌株的单株选优和通过栽培管理等途径提高种核的品质和产量提供了理论依据。
陈鹏. 1991. 目前国内外银杏研究进展概况. 浙江林业科技, 11(4): 70-75;69. |
陈鹏, 何凤仁, 余碧钰, 等. 1994. 60Co和速冻对银杏种胚及氢氰酸含量的影响. 江苏农学院学报, 15(1): 53-56. |
陈鹏, 何凤仁, 褚生华, 等. 1996. 银杏早果丰产的理论基础及其栽培技术. 果树科学, 13(4): 255-256. |
陈鹏, 何凤仁, 褚生华, 等. 1997. 银杏种实丰产单株选优研究. 园艺学报, 24(2): 205-207. DOI:10.3321/j.issn:0513-353X.1997.02.025 |
陈鹏, 何凤仁, 褚生华等. 银杏品种选优及其结构调整. 见: 全国第八次银杏学术研讨会论文集. 武汉: 湖北科学技术出版社, 2000: 74-91
|
陈鹏. 银杏集约化栽培及其产业化技术开发的研究进展. 见: 全国第十次银杏学术研讨会论文集. 北京: 中国农业科学技术出版社, 2002: 65-76
|
傅德志, 杨亲二. 1993. 银杏雌性生殖器官的形态学本质及其系统学意义. 植物分类学报, 31(3): 294-296. |
郭善基主编. 中国果树志•银杏卷. 北京: 中国林业出版社, 1993: 78-91
|
何凤仁. 1989. 银杏的栽培. 南京: 江苏科学技术出版社.
|
吉成均. 1999. 银杏大孢子形成的超微结构研究. 植物学报, 41(12): 1323-1326. DOI:10.3321/j.issn:1672-9072.1999.12.014 |
莫惠栋. 1992. 农业试验统计. 第二版. 上海: 上海科技出版社.
|
陶俊, 陈鹏. 1998. 银杏传粉对雌花生长及过氧化物酶同工酶的影响. 江苏农学院学报, 19(3): 53-56. |
王伏雄, 陈祖铿. 1983. 银杏胚胎发育的研究——兼论银杏目的亲缘关系. 植物学报, 25(3): 199-207. |
曾勉. 1935. 浙江诸暨之银杏. 园艺, 1(5): 157-165. |
Foster A S, Gifford E M. 李正理等译. 维管植物比较形态学. 北京: 科学出版社, 1983: 362-378
|
Chen P, He F R, Yu B Y, et al. 1999. Seed stone shape and the relative component in kernel of Ginkgo biloba. Forestry Studies in China, 1(1): 42-47. |
Davidson K G V. 1994. Comparison of seed-quality indices resulting from single-seed electroconductivity measurements. Hortscience, 29(10): 1158-1163. DOI:10.21273/HORTSCI.29.10.1158 |
Friedman W E. 1987. Morphogenesis and experimental aspects of growth and development of the male gametpohyte of Ginkgo biloba in vitro. Amer J Bot, 74(12): 1816-1831. DOI:10.1002/j.1537-2197.1987.tb08784.x |
Friedman W E, Gifford E M. 1988. Division of the generative cell and late development in the male gametophyte of Ginkgo biloba. Amer J Bot, 75(9): 1434-1442. DOI:10.1002/j.1537-2197.1988.tb14203.x |